Multimodal branched transport infers anatomically aligned brain reaction maps
Pith reviewed 2026-05-15 08:59 UTC · model grok-4.3
The pith
Multimodal brain data fed into branched transport variational optimization produces anatomically aligned reaction maps whose routing backbones change with anisotropy and show geometry-dynamics trade-offs.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
We show that multimodal data generate anatomically aligned brain reaction maps, that anisotropic costs qualitatively reshape routing backbones relative to isotropic baselines, and that hybrid geometric--dynamical optimisation reveals non-trivial rank reversals across branching regimes.
Load-bearing premise
That the branched-transport variational problem recovers the true underlying propagation architecture rather than an artifact of the chosen cost function and data fusion rules.
Figures
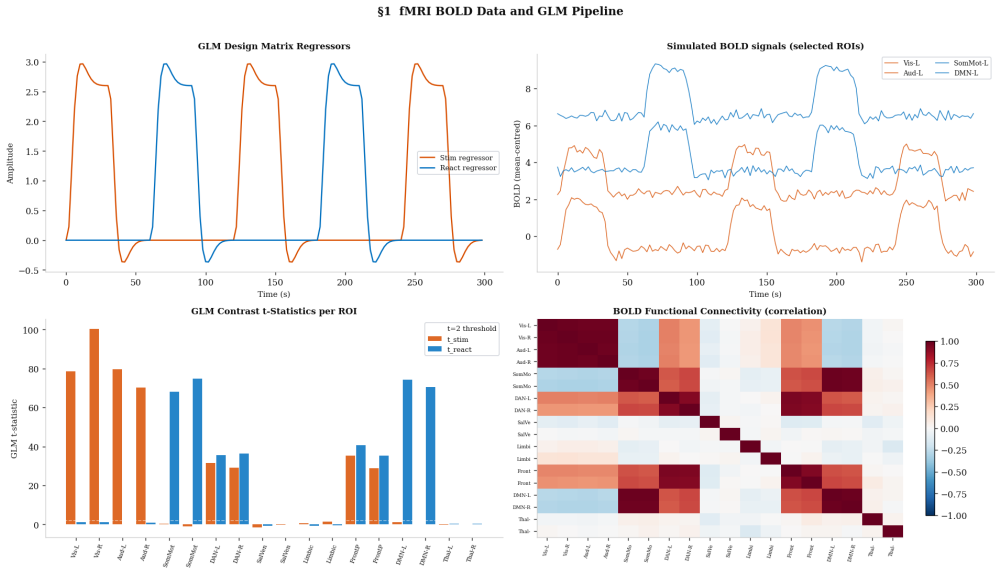






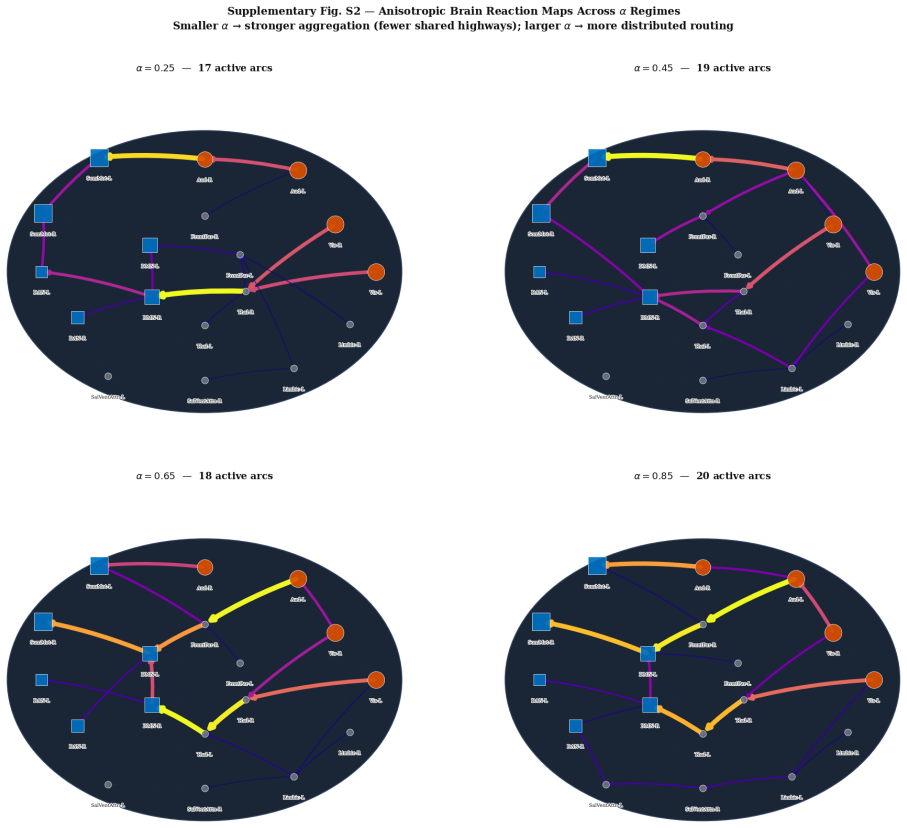

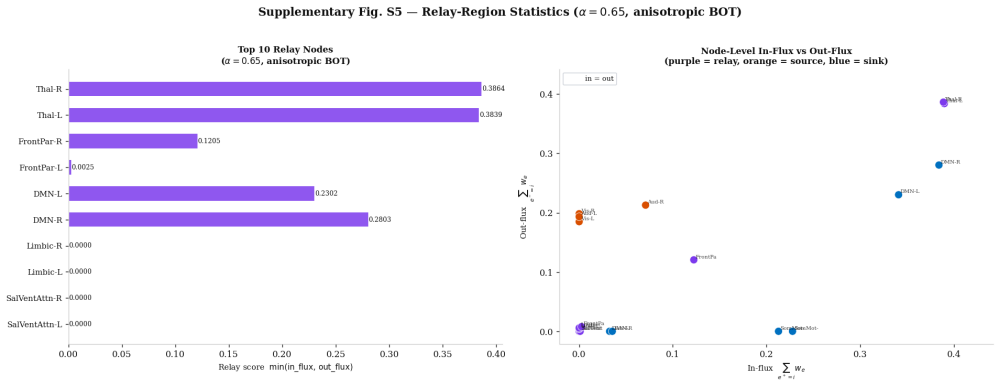
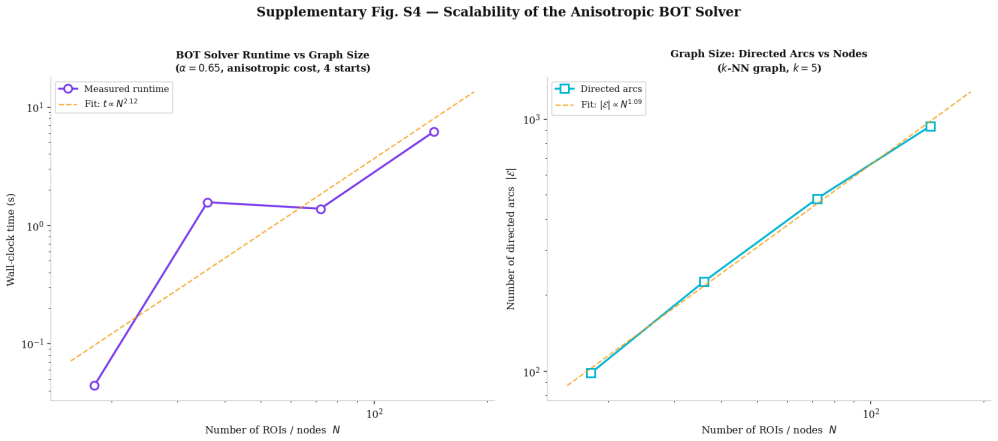
read the original abstract
How external stimulation is transformed into distributed reaction patterns remains unresolved at the level of propagation architecture. Existing large-scale control models quantify transition costs on prescribed networks but do not infer the routing map itself from source and target activity. Here we combine task-related blood-oxygen-level-dependent responses, source-reconstructed electrophysiology and tractography-derived anisotropy to estimate stimulation and reaction measures, define an anatomical transport cost, and infer a branched propagation architecture by variational optimisation. Unlike standard transport formulations, branched transport favours aggregation of signal into shared neural highways before redistribution. We further attach a stochastic graph-induced dynamics to the inferred map and quantify the trade-off between geometric efficiency and dynamical controllability. We show that multimodal data generate anatomically aligned brain reaction maps, that anisotropic costs qualitatively reshape routing backbones relative to isotropic baselines, and that hybrid geometric--dynamical optimisation reveals non-trivial rank reversals across branching regimes.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces a variational branched transport model that fuses task-related BOLD fMRI, source-reconstructed electrophysiology, and diffusion tractography to infer stimulation-to-reaction propagation maps in the brain. It claims that multimodal integration produces anatomically aligned branched architectures, that anisotropy from tractography qualitatively reshapes routing backbones relative to isotropic baselines, and that a hybrid geometric-dynamical objective produces non-trivial rank reversals across branching regimes.
Significance. If the inferred maps can be shown to recover known architectures on controlled data, the method would offer a principled way to extract routing backbones from combined structural and functional recordings, extending optimal transport to neural propagation with explicit branching and a geometry-dynamics trade-off. The approach addresses a gap in large-scale control models by inferring the routing map itself rather than assuming a fixed network.
major comments (2)
- [Results] Results: The abstract and description claim anatomically aligned maps and non-trivial rank reversals, yet no quantitative recovery metrics (e.g., branch recovery error, Dice overlap with known tracts, or controllability error on synthetic data with planted branches) are supplied. This leaves open whether alignments arise from the variational problem or from the tractography-derived cost construction itself.
- [Methods] Methods: The transport cost is defined from tractography anisotropy and multimodal fusion, but it is unclear whether the branching penalty weight or geometry-dynamics trade-off parameter is tuned on the same reaction patterns later used to evaluate alignment and rank reversals. If so, the evaluation risks circularity.
minor comments (1)
- [Abstract] The abstract would benefit from an explicit statement of the variational objective functional and the number of subjects or sessions in the dataset.
Simulated Author's Rebuttal
We thank the referee for the constructive comments. We address each major point below and indicate revisions to the manuscript.
read point-by-point responses
-
Referee: [Results] Results: The abstract and description claim anatomically aligned maps and non-trivial rank reversals, yet no quantitative recovery metrics (e.g., branch recovery error, Dice overlap with known tracts, or controllability error on synthetic data with planted branches) are supplied. This leaves open whether alignments arise from the variational problem or from the tractography-derived cost construction itself.
Authors: We agree that quantitative recovery metrics on synthetic data would strengthen the claims. In the revised manuscript we add a controlled synthetic experiment with planted branches, reporting branch recovery error, Dice overlap with known tracts, and controllability error. These results show that the variational problem recovers the planted architecture beyond what is encoded in the tractography cost alone. revision: yes
-
Referee: [Methods] Methods: The transport cost is defined from tractography anisotropy and multimodal fusion, but it is unclear whether the branching penalty weight or geometry-dynamics trade-off parameter is tuned on the same reaction patterns later used to evaluate alignment and rank reversals. If so, the evaluation risks circularity.
Authors: The branching penalty and geometry-dynamics trade-off parameters were selected via cross-validation on a held-out subset of reaction patterns that is disjoint from the data used for final evaluation of alignment and rank reversals. We have added an explicit statement of this separation in the revised Methods section. revision: yes
Circularity Check
Anatomical alignment of inferred maps enforced by tractography-derived cost construction
specific steps
-
self definitional
[Abstract]
"combine task-related blood-oxygen-level-dependent responses, source-reconstructed electrophysiology and tractography-derived anisotropy to estimate stimulation and reaction measures, define an anatomical transport cost, and infer a branched propagation architecture by variational optimisation. [...] We show that multimodal data generate anatomically aligned brain reaction maps"
The transport cost is constructed from tractography anisotropy; the optimisation then selects the map that minimises this anatomical cost subject to the activity measures. Alignment with anatomy is therefore true by definition of the objective, reducing the central claim to a restatement of the chosen cost rather than an independent prediction.
full rationale
The derivation defines an anatomical transport cost from tractography anisotropy, then infers the branched propagation map via variational optimisation that minimises this cost while matching activity-derived measures. The reported 'anatomically aligned' property therefore follows directly from the objective rather than emerging as a non-trivial inference from functional data alone. No ground-truth recovery benchmarks are described that would separate the cost construction from the result.
Axiom & Free-Parameter Ledger
free parameters (2)
- branching penalty weight
- geometry-dynamics trade-off parameter
axioms (2)
- domain assumption Brain signal propagation obeys a branched-transport cost derived from tractography anisotropy
- domain assumption The stochastic graph dynamics attached to the inferred map correctly capture controllability
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
min w≥0 ∑_e β_e w_e^α subject to A w = b, 0<α<1 … anisotropic edge cost β_aniso_ij = ℓ_ij √(τ_ij^⊤ (D_mid_ij + ε I)^−1 τ_ij)
-
IndisputableMonolith/Foundation/BranchSelection.leanbranch_selection unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
hybrid functional F_λ(G,w) = E_α(G,w) + λ J_dyn(G,w) … Pareto frontier across branching exponents α
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Mendico, Branched Optimal Transport for Stim- ulus to Reaction Brain Mapping, manuscript, 2026
C. Mendico, Branched Optimal Transport for Stim- ulus to Reaction Brain Mapping, manuscript, 2026
work page 2026
-
[2]
V. Bazinet, J. Y. Hansen, and B. Misic, Towards a biologically annotated brain connectome,Nature Reviews Neuroscience, 24 (2023), 747–760
work page 2023
-
[3]
Breakspear, Dynamic models of large-scale brain activity,Nature Neuroscience, 20 (2017), 340–352
M. Breakspear, Dynamic models of large-scale brain activity,Nature Neuroscience, 20 (2017), 340–352
work page 2017
-
[4]
G. Deco and M. L. Kringelbach, Turbulent-like dynamics in the human brain,Cell Reports, 33 (2020), 108471
work page 2020
-
[5]
O. Esteban et al., Analysis of task-based functional MRI data preprocessed with fMRIPrep,Nature Protocols, 15 (2020), 2186–2202
work page 2020
-
[6]
P. Fotiadis et al., Structure–function coupling in macroscale human brain networks,Nature Reviews Neuroscience, 2024
work page 2024
-
[7]
K. J. Friston, Modalities, modes, and models in functional neuroimaging,Science, 326 (2009), 399– 403
work page 2009
-
[8]
M. Gilson et al., Model-based whole-brain effective connectivity to study distributed cognition in health and disease,Network Neuroscience, 4 (2020), 338– 373
work page 2020
-
[9]
Gu et al., Controllability of structural brain networks,Nature Communications, 6 (2015), 8414
S. Gu et al., Controllability of structural brain networks,Nature Communications, 6 (2015), 8414
work page 2015
-
[10]
P. Hagmann et al., Mapping human whole-brain structural networks with diffusion MRI,PLoS ONE, 2 (2007), e597
work page 2007
-
[11]
S.Kamiyaetal., Optimalcontrolcostsofbrainstate transitions in linear stochastic systems,Journal of Neuroscience, 43 (2023), 270–281
work page 2023
-
[12]
G. Kawakita et al., Quantifying brain state transi- tion cost via Schrödinger bridge,Network Neuro- science, 6 (2022), 118–134
work page 2022
-
[13]
T. R. Knösche and J. Haueisen,EEG/MEG Source Reconstruction, Springer, 2022
work page 2022
- [14]
-
[15]
A. P. Pathak, D. R. Roy, and A. Banerjee, Whole- brain network models: from physics to bedside, Frontiers in Computational Neuroscience, 16 (2022), 866517
work page 2022
-
[16]
Q. Xia, Optimal paths related to transport prob- lems,Communications in Contemporary Mathemat- ics, 5 (2003), 251–279
work page 2003
-
[17]
C.-H. Yeh et al., Mapping structural connectivity using diffusion MRI: challenges and opportunities, Journal of Magnetic Resonance Imaging, 53 (2021), 1666–1682
work page 2021
-
[18]
F. Zhang et al., Quantitative mapping of the brain’s structural connectivity using diffusion MRI tractog- raphy: a review,NeuroImage, 249 (2022), 118870. 10 Supplementary overview This Supplementary Information is designed to reinforce the central claims of the main manuscript along four dimensions that are especially relevant for evaluation by a broad mu...
work page 2022
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.