Collective spatial reorganization from arrest to peeling and migration through density-dependent mobility in internal-state coordinates
Pith reviewed 2026-05-10 18:50 UTC · model grok-4.3
The pith
Mobility in an internal-state coordinate alone drives particle aggregates from arrest to peeling migration
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
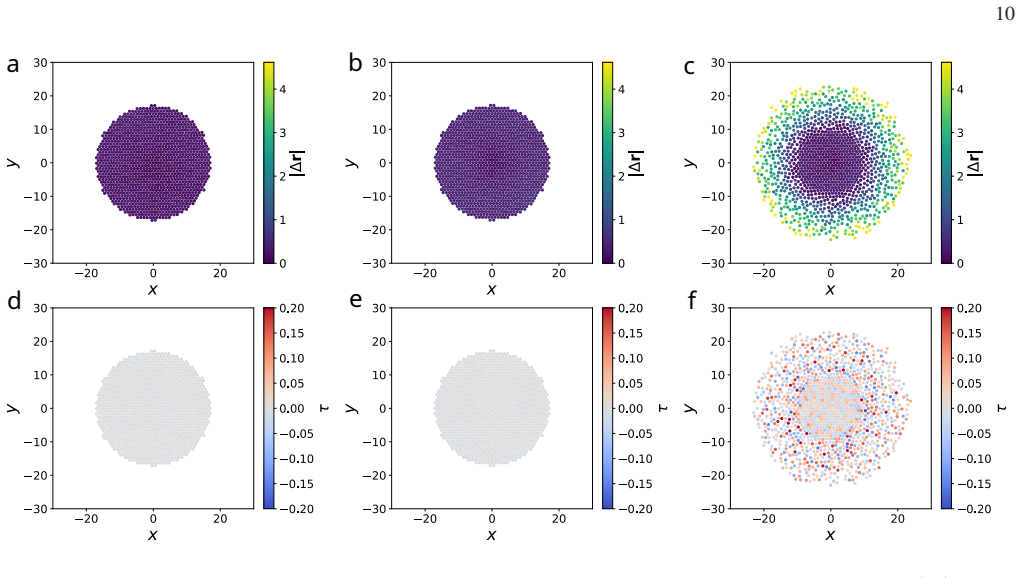
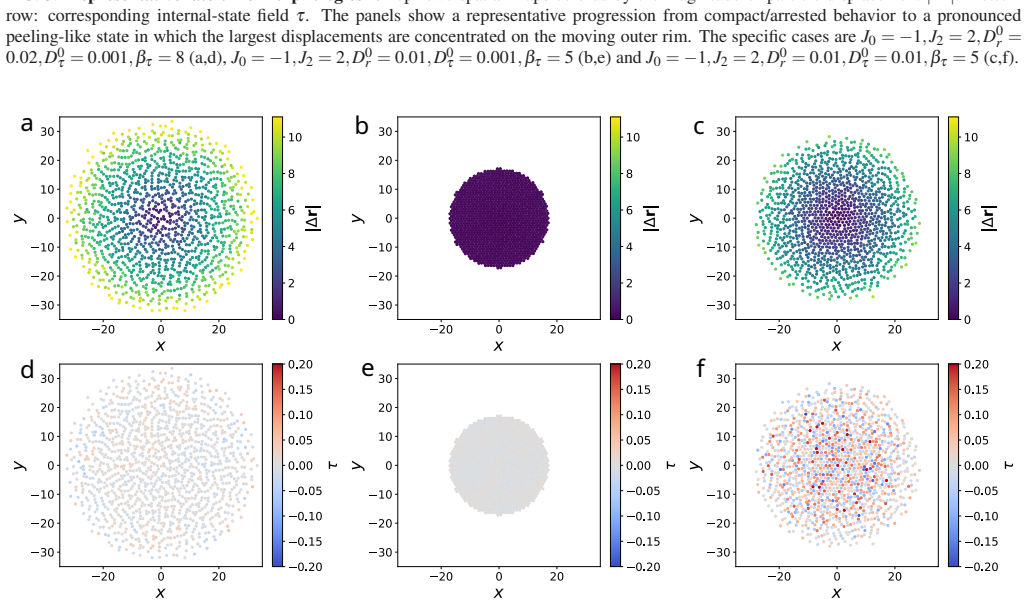
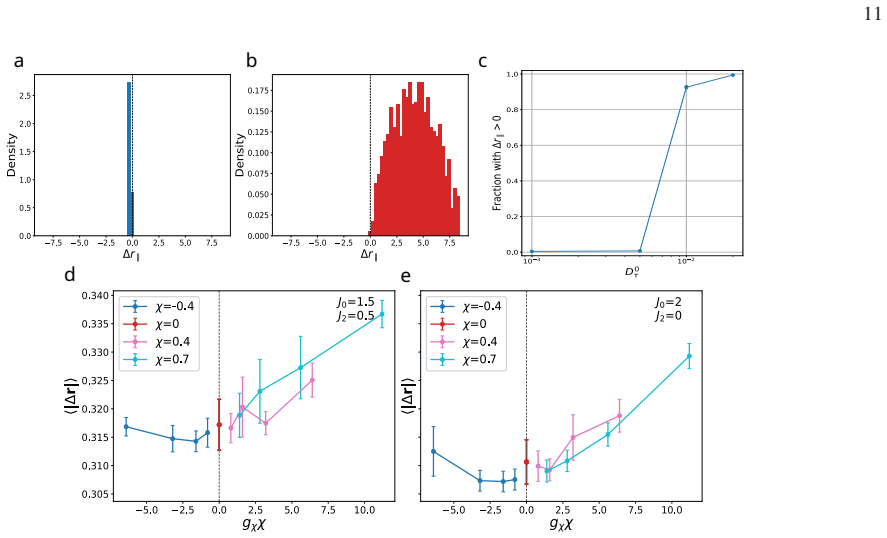
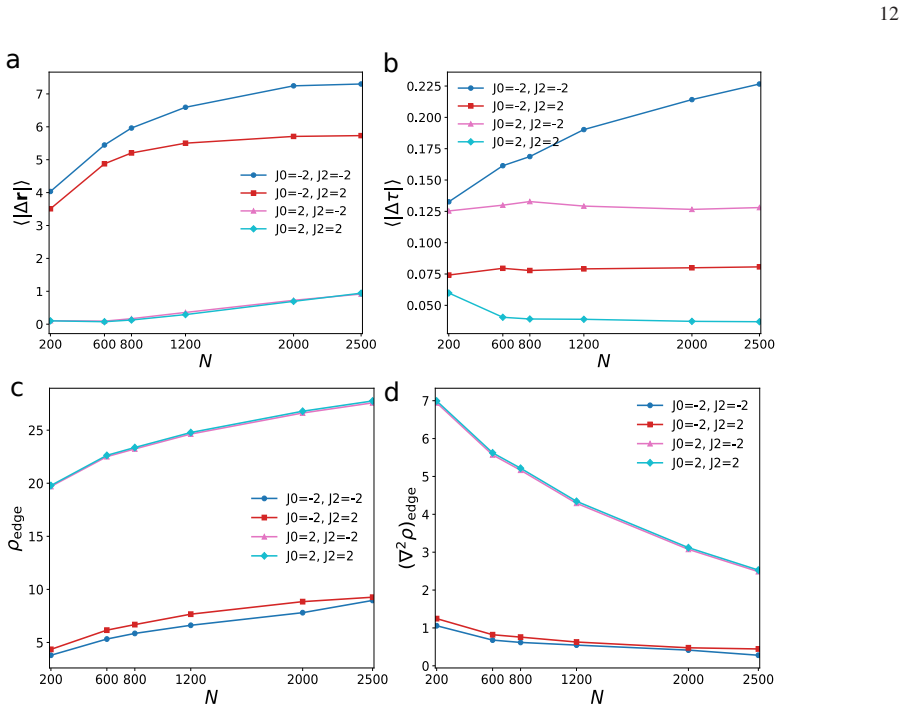
Mobility in internal-state coordinates alone provides an independent control axis for large-scale spatial reorganization. Increasing the baseline internal-state diffusivity and tuning its density dependence drives a transition from arrested aggregates to a peeling-like regime with broad spatial excursions, strong outward radial bias, and edge-localized activity, while the baseline positional diffusivity is held fixed. The transition is accompanied by correlated broadening of spatial and internal-state displacements, systematic reorganization of radial density and density-curvature profiles, and a pronounced dependence on system size, consistent with the idea that growing aggregates can cross
What carries the argument
Asymmetrically cross-coupled density-dependent mobilities defined on both spatial and internal-state coordinates within an overdamped stochastic particle model.
If this is right
- The transition is accompanied by correlated broadening of spatial and internal-state displacements.
- Radial density and density-curvature profiles reorganize systematically.
- The behavior exhibits pronounced dependence on system size, consistent with growing aggregates crossing into a boundary-dominated migratory state.
- State change, spatial redistribution, and neighborhood structure can be modeled within a common formalism.
Where Pith is reading between the lines
- The size dependence implies that sufficiently large aggregates are more likely to exhibit the migratory regime, which could be tested by systematically varying system size in further simulations.
- The same control axis might be relevant to other crowded systems in which internal states modulate effective mobility, such as active-matter or tissue models.
- The unified formalism motivates extending the approach to include neighborhood structure evolution alongside spatial and state dynamics.
Load-bearing premise
The model assumes finite-range forces, asymmetrically cross-coupled density-dependent mobilities on spatial and internal-state coordinates, and the overdamped limit for deriving the stochastic update equations.
What would settle it
A simulation in which internal-state diffusivity is increased and its density dependence is tuned yet no transition to outward-biased peeling or edge activity appears would falsify the claim that internal-state mobility supplies an independent control axis.
Figures

read the original abstract
Numerous problems in development, regeneration, and disease involve simultaneous evolution of both spatial organization and the internal state of the constituents in addition to local interactions and crowding. This motivates us to study a minimal model for interacting populations evolving in coupled spatial and internal-state coordinates. We focus on a specific transition of particular biological interest: the reorganization of dense collectives from compact or arrested states toward boundary-led peeling and migration. In our formulation, each particle carries a spatial position and a scalar internal state, and interacts through finite-range forces. Mobilities are defined on both spatial and internal-state coordinates with a density dependence, and are asymmetrically cross-coupled. We derive update equations for stochastic dynamics in the overdamped limit and perform numerical simulations. We find that mobility in internal-state coordinates alone provides an independent control axis for large-scale spatial reorganization. In particular, increasing the baseline internal-state diffusivity and tuning its density dependence drives a transition from arrested aggregates to a peeling-like regime with broad spatial excursions, strong outward radial bias, and edge-localized activity, while the baseline positional diffusivity is held fixed. The transition is accompanied by correlated broadening of spatial and internal-state displacements, systematic reorganization of radial density and density-curvature profiles, and a pronounced dependence on system size, consistent with the idea that growing aggregates can cross into a boundary-dominated migratory state. These results establish the utility of our approach and motivate a broader framework aimed at modeling state change, spatial redistribution, and neighborhood structure within a common formalism.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces a minimal model of interacting particles evolving in coupled spatial and internal-state coordinates, with finite-range forces and asymmetrically cross-coupled, density-dependent mobilities on both coordinates. Overdamped stochastic dynamics are derived and simulated numerically. The central claim is that mobility in the internal-state coordinate alone acts as an independent control axis: increasing its baseline diffusivity and tuning its density dependence drives a transition from arrested aggregates to a peeling-like regime featuring broad spatial excursions, strong outward radial bias, and edge-localized activity, while the baseline positional diffusivity remains fixed. The transition is accompanied by correlated broadening of displacements, reorganization of radial density and curvature profiles, and pronounced system-size dependence.
Significance. If the independence of the internal-state control axis is rigorously established, the work offers a compact framework for linking internal-state dynamics to large-scale spatial reorganization in dense collectives. This has potential relevance to biological processes involving collective migration and boundary-driven transitions. The direct simulation of the stated model rules, without reliance on fitted quantities or self-referential derivations, is a positive feature that supports falsifiability.
major comments (2)
- [Model derivation and simulation setup] The claim that internal-state mobility provides an 'independent control axis' while 'the baseline positional diffusivity is held fixed' is load-bearing for the central result. Given the asymmetric cross-coupling of the mobilities, it is unclear whether changes to the internal-state baseline diffusivity D_s indirectly modify the effective positional mobility through the coupling terms. The model definition (in the section deriving the stochastic update equations) should explicitly state the functional forms of the mobility matrix elements and include a direct verification, such as low-density mean-squared displacement measurements, confirming that the positional diffusion coefficient remains unchanged when D_s is varied.
- [Numerical results] The abstract states that numerical simulations support the transition and accompanying profile changes, yet no information is supplied on the specific parameter values, system sizes, number of realizations, error bars, or controls for finite-size effects. Without these, the quantitative robustness of the reported radial bias, edge activity, and system-size dependence cannot be assessed, weakening the evidential basis for the transition.
minor comments (2)
- [Abstract] The abstract is information-dense; splitting the description of the transition and its signatures into shorter sentences would improve readability.
- [Model section] Notation for the mobilities and cross-coupling terms should be introduced with a clear table or explicit equations early in the model section to aid the reader in following the asymmetric coupling.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments on our manuscript. We address each major comment below and will revise the manuscript to improve clarity, completeness, and reproducibility.
read point-by-point responses
-
Referee: [Model derivation and simulation setup] The claim that internal-state mobility provides an 'independent control axis' while 'the baseline positional diffusivity is held fixed' is load-bearing for the central result. Given the asymmetric cross-coupling of the mobilities, it is unclear whether changes to the internal-state baseline diffusivity D_s indirectly modify the effective positional mobility through the coupling terms. The model definition (in the section deriving the stochastic update equations) should explicitly state the functional forms of the mobility matrix elements and include a direct verification, such as low-density mean-squared displacement measurements, confirming that the positional diffusion coefficient remains unchanged when D_s is varied.
Authors: We agree that explicit documentation of the mobility matrix and a direct verification are necessary to substantiate the independence of the internal-state control axis. In the revised manuscript, we will expand the model derivation section to state the functional forms of all mobility matrix elements, including the asymmetric cross-coupling. The baseline positional diffusivity D_x enters only through its own diagonal term (multiplied by a density-dependent factor independent of D_s), while D_s enters only through its diagonal term (with its own density dependence); the off-diagonal cross terms couple the coordinates via forces but do not mix the baseline diffusivities. We will also add low-density mean-squared displacement measurements (in the dilute limit where interactions vanish) demonstrating that the effective positional diffusion coefficient remains equal to the fixed D_x value across the range of D_s explored. These additions will rigorously confirm that internal-state mobility acts as an independent control axis. revision: yes
-
Referee: [Numerical results] The abstract states that numerical simulations support the transition and accompanying profile changes, yet no information is supplied on the specific parameter values, system sizes, number of realizations, error bars, or controls for finite-size effects. Without these, the quantitative robustness of the reported radial bias, edge activity, and system-size dependence cannot be assessed, weakening the evidential basis for the transition.
Authors: We thank the referee for identifying this omission, which affects the ability to assess quantitative robustness. In the revised manuscript, we will add a dedicated 'Simulation parameters and controls' subsection (or supplementary table) that reports: all fixed and varied parameter values (including D_x, the range of D_s, density-dependence exponents, interaction cutoff, temperature, and timestep); system sizes (particle number N and domain dimensions); number of independent realizations (ensemble size); error bars (standard errors computed over realizations); and finite-size controls (explicit comparisons across multiple N values confirming the reported system-size dependence of the transition). These details will enable readers to evaluate the statistical reliability of the radial bias, edge activity, and scaling behavior. revision: yes
Circularity Check
No circularity: results follow from direct simulation of explicitly stated model rules with no reductions to inputs or self-citations.
full rationale
The paper defines a minimal interacting-particle model with finite-range forces, density-dependent mobilities on spatial and internal-state coordinates, and asymmetric cross-coupling. It derives the overdamped stochastic update equations from this setup and obtains all reported transitions (arrested aggregates to peeling/migration) via numerical simulation while claiming to hold the baseline positional diffusivity fixed. No equations or results are shown to reduce by construction to fitted parameters, prior self-citations, or ansatzes imported from the authors' own work; the observed radial bias, edge activity, and system-size dependence emerge as simulation outputs rather than tautological restatements of the input rules. The skeptic's concern about effective positional mobility under cross-coupling addresses model validity and independence of control axes, not circularity in the derivation chain itself. This is the standard non-circular case for a simulation-based study.
Axiom & Free-Parameter Ledger
free parameters (2)
- baseline internal-state diffusivity
- density dependence of internal-state mobility
axioms (3)
- domain assumption Overdamped limit applies to the stochastic dynamics
- domain assumption Interactions occur via finite-range forces
- ad hoc to paper Mobilities are asymmetrically cross-coupled and density-dependent
Reference graph
Works this paper leans on
-
[1]
Quantitative approaches in developmental biology
Andrew C Oates, Nicole Gorfinkiel, Marcos Gonzalez-Gait an, and Carl-Philipp Heisenberg. Quantitative approaches in developmental biology. Nature Reviews Genetics, 10(8):517–530, 2009
work page 2009
-
[2]
Towards a physical und erstanding of developmental patterning
Jose Negrete Jr and Andrew C Oates. Towards a physical und erstanding of developmental patterning. Nature Reviews Genetics , 22(8):518–531, 2021
work page 2021
-
[3]
Y ue Liu, Xufeng Xue, Shiyu Sun, Norio Kobayashi, Y ung Su K im, and Jianping Fu. Morphogenesis beyond in vivo. Nature Reviews Physics, 6(1):28–44, 2024
work page 2024
-
[4]
Spatial biology of cancer evolution
Zaira Seferbekova, Artem Lomakin, Lucy R Yates, and Mori tz Gerstung. Spatial biology of cancer evolution. Nature Reviews Genetics, 24(5):295–313, 2023
work page 2023
-
[5]
Spatial self-o rganization of metabolism in microbial systems: a matter of enzymes and chemicals
Martin Ackermann, Simon van Vliet, et al. Spatial self-o rganization of metabolism in microbial systems: a matter of enzymes and chemicals. Cell systems, 14(2):98–108, 2023
work page 2023
-
[6]
Image-b ased modelling of organogenesis
Dagmar Iber, Zahra Karimaddini, and Erkan Ünal. Image-b ased modelling of organogenesis. Briefings in Bioinformatics , 17(4):616–627, 2016
work page 2016
-
[7]
Dario Bressan, Giorgia Battistoni, and Gregory J Hannon . The dawn of spatial omics. Science, 381(6657):eabq4964, 2023
work page 2023
-
[8]
Principles and challen ges of modeling temporal and spatial omics data
Britta V elten and Oliver Stegle. Principles and challen ges of modeling temporal and spatial omics data. Nature Methods, 20(10):1462– 1474, 2023
work page 2023
-
[9]
Establishing a co nceptual framework for holistic cell states and state trans itions
Susanne M Rafelski and Julie A Theriot. Establishing a co nceptual framework for holistic cell states and state trans itions. Cell, 187(11):2633–2651, 2024
work page 2024
-
[10]
Peijie Zhou, Shuxiong Wang, Tiejun Li, and Qing Nie. Dis secting transition cells from single-cell transcriptome d ata through multiscale stochastic dynamics. Nature communications, 12(1):5609, 2021
work page 2021
-
[11]
Cellrank for directe d single-cell fate mapping
Marius Lange, V olker Bergen, Michal Klein, Manu Setty, Bernhard Reuter, Mostafa Bakhti, Heiko Lickert, Meshal Ans ari, Janine Schniering, Herbert B Schiller, et al. Cellrank for directe d single-cell fate mapping. Nature methods, 19(2):159–170, 2022
work page 2022
-
[12]
Optimal transport for singl e-cell and spatial omics
Charlotte Bunne, Geoffrey Schiebinger, Andreas Kraus e, Aviv Regev, and Marco Cuturi. Optimal transport for singl e-cell and spatial omics. Nature Reviews Methods Primers, 4(1):58, 2024
work page 2024
-
[13]
Self-organization of stem cells into embryos: a win dow on early mammalian development
Marta N Shahbazi, Eric D Siggia, and Magdalena Zernicka -Goetz. Self-organization of stem cells into embryos: a win dow on early mammalian development. Science, 364(6444):948–951, 2019
work page 2019
-
[14]
Theory of branching morphogenesis by loca l interactions and global guidance
Mehmet Can Uçar, Dmitrii Kamenev, Kazunori Sunadome, D ominik Fachet, Francois Lallemend, Igor Adameyko, Saida Ha djab, and Edouard Hannezo. Theory of branching morphogenesis by loca l interactions and global guidance. Nature Communications, 12(1):6830, 2021
work page 2021
-
[15]
A method to recapitulate early embryoni c spatial patterning in human embryonic stem cells
Aryeh Warmflash, Benoit Sorre, Fred Etoc, Eric D Siggia, and Ali H Brivanlou. A method to recapitulate early embryoni c spatial patterning in human embryonic stem cells. Nature methods, 11(8):847–854, 2014
work page 2014
-
[16]
Single-cell tracking reveals super -spreading brain cancer cells with high persistence
Aimilia Nousi, Maria Tangen Søgaard, Mélanie Audoin, a nd Liselotte Jauffred. Single-cell tracking reveals super -spreading brain cancer cells with high persistence. Biochemistry and Biophysics Reports , 28:101120, 2021
work page 2021
-
[17]
Statistic al mechanics meets single-cell biology
Andrew E Teschendorff and Andrew P Feinberg. Statistic al mechanics meets single-cell biology. Nature Reviews Genetics, 22(7):459– 476, 2021
work page 2021
-
[18]
Kinetic reconstru ction of free energies as a function of multiple order parame ters
Yagyik Goswami and Srikanth Sastry. Kinetic reconstru ction of free energies as a function of multiple order parame ters. The Journal of Chemical Physics, 158(14), 2023
work page 2023
-
[19]
Ligang Zhu and Jin Wang. Quantifying landscape and flux f rom single-cell omics: unraveling the physical mechanisms of cell function. JACS Au, 5(8):3738–3757, 2025
work page 2025
-
[20]
Hydrodynamic theory of active matter
Frank Jülicher, Stephan W Grill, and Guillaume Salbreu x. Hydrodynamic theory of active matter. Reports on Progress in Physics , 81(7):076601, 2018
work page 2018
-
[21]
Computational models for activ e matter
M Reza Shaebani, Adam Wysocki, Roland G Winkler, Gerhar d Gompper, and Heiko Rieger. Computational models for activ e matter. Nature Reviews Physics, 2(4):181–199, 2020
work page 2020
-
[22]
Yielding behaviour of active particles in bulk and in confine ment
Yagyik Goswami, GV Shivashankar, and Srikanth Sastry. Yielding behaviour of active particles in bulk and in confine ment. Nature Physics, pages 1–8, 2025
work page 2025
-
[23]
Activity-induc ed annealing leads to a ductile-to-brittle transition in am orphous solids
Rishabh Sharma and Smarajit Karmakar. Activity-induc ed annealing leads to a ductile-to-brittle transition in am orphous solids. Nature Physics, pages 1–9, 2025
work page 2025
-
[24]
Stochastic processes in physics and chemistry
NG V an Kampen. Stochastic processes in physics and chemistry . Elsevier B.V ., third edition, 2007
work page 2007
-
[25]
Oskar Hallatschek, Sujit S Datta, Knut Drescher, Jörn D unkel, Jens Elgeti, Bartek Waclaw, and Ned S Wingreen. Proli ferating active matter. Nature Reviews Physics, 5(7):407–419, 2023
work page 2023
-
[26]
Physical models of coll ective cell migration
Ricard Alert and Xavier Trepat. Physical models of coll ective cell migration. Annual Review of Condensed Matter Physics, 11(1):77–101, 2020
work page 2020
-
[27]
Sel f-aligning polar active matter
Paul Baconnier, Olivier Dauchot, Vincent Démery, Gust avo Düring, Silke Henkes, Cristián Huepe, and Amir Shee. Sel f-aligning polar active matter. Reviews of Modern Physics , 97(1):015007, 2025
work page 2025
-
[28]
Growth, homeo static regulation and stem cell dynamics in tissues
E Hannezo, Jacques Prost, and J-F Joanny. Growth, homeo static regulation and stem cell dynamics in tissues. Journal of The Royal Society Interface, 11(93), 2014. 14 APPENDIX Appendix A: From the Hamiltonian to a continuity equation in shared coordinates
work page 2014
-
[29]
Empirical density and continuity equation Let the empirical density in the joint space of position and t ype be ˆρ (r,τ ,t) = N ∑ i=1 δ ( r − ri(t) ) δ ( τ − τ i(t) ) . (A1) Differentiating with respect to time gives ∂ t ˆρ = − ∑ i ˙ri ·∇ r [ δ ( r − ri ) δ ( τ − τ i ) ] − ∑ i ˙τ i ∂ τ [ δ ( r − ri ) δ ( τ − τ i ) ] = − ∇ r·∑ i ˙riδ ( r − ri ) δ ( τ − τ i...
-
[30]
Free-energy functional associated with the microscopic Hamiltonian Starting from the broad Hamiltonian H = ∑ i< j U(ri − r j,τ i,τ j) +∑ i Vintr(τ i), (A5) the corresponding mean-field free-energy functional is F [ρ ] = kBT ∫ d2r dτ ρ (r,τ ) [ ln ρ (r,τ ) − 1 ] + 1 2 ∫ d2r dτ d2r′dτ ′ρ (r,τ )U(r − r′,τ ,τ ′) ρ (r′,τ ′) + ∫ d2r dτ ρ (r,τ )Vintr(τ ). (A6) Th...
-
[31]
Continuum currents and Onsager form At the continuum level, conservation in the shared coordina tes takes the form ∂ t ρ = − ∇ r·Jr − ∂ τ Jτ . (A11) This corresponds to the following block mobility equation ( Jr Jτ ) = − ρ ( Mrr Mrτ Mτ r Mττ ) ( ∇ rµ ∂ τ µ ) , (A12) where Mrr is a 2 × 2 mobility block, Mττ is a scalar mobility in type space, and the off-d...
-
[32]
Connection to the microscopic equations used in this work The particle dynamics can be written as ( ˙ri ˙τ i ) = ( Mrr(ρ i) Mrτ (ρ i) Mτ r(ρ i) Mττ (ρ i) ) ( F(r) i F (τ ) i ) + ξnoise i , (A14) with F(r) i = − ∂ H ∂ ri , F (τ ) i = − ∂ H ∂ τ i . (A15) For the exact implementation used in the present manuscript , the conservative force sector is given by ...
-
[33]
Covariance matrix for one particle For one particle in two spatial dimensions, define the stocha stic increment vector ∆ Wi = η i,x η i,y ζ i . (A1) In the most general, equilibrium case, the target covarianc e over one Euler–Maruyama step is ⟨ ∆ Wi ∆ WT i ⟩ = Σ i = 2kBT ∆ t Mr( ρ i) 0 M× (ρ i) 0 Mr(ρ i) M× (ρ i) M× (ρ i) M× (ρ i) Mτ (ρ i) ...
-
[34]
(A2) is positive semidefinite only if 2M× (ρ i)2 ≤ Mr(ρ i)Mτ (ρ i)
Positive-semidefinite constraint and numerical draw The 3 × 3 covariance matrix in Eq. (A2) is positive semidefinite only if 2M× (ρ i)2 ≤ Mr(ρ i)Mτ (ρ i). (A6) In practice, the implementation enforces this by clamping |M× |to a value just below the bound, |M× | ≤ 0.999 √ MrMτ 2 . (A7) Once the covariance matrix is constructed, the increment ca n be sampled ...
-
[35]
(A2), th en one recovers the standard overdamped fluctuation–dissip ation structure
Detailed balance: benchmark and caveat If the mobility matrix is reciprocal, the deterministic dri ft is derived from a common chemical potential, and the noise co- variance is matched to the same mobility through Eq. (A2), th en one recovers the standard overdamped fluctuation–dissip ation structure. In the simplest constant-mobility case this yie lds an ...
-
[36]
Reciprocal passive benchmark The first reference description is the reciprocal passive mo del. Here the currents in the shared coordinates are generat ed by a single chemical potential µ and a reciprocal block mobility, Mrτ = MT τ r, (A1) with covariance-consistent noise. In this limit the spatia l and type sectors are coupled, but the coupling is still co...
-
[37]
Density-dependent mobility without explicit active cro ss-drift A second layer of complexity arises when the mobilities depe nd on local density, Mr = Mr(ρ ), Mτ = Mτ (ρ ), M× = M× (ρ ). (A2) Even before any explicit active asymmetry is introduced, th is already creates a bulk–rim contrast in transport: dense r egions are dynamically slowed relative to di...
-
[38]
Asymmetric active cross-drift used in the present work The production model used for the figures departs more strong ly from equilibrium. The deterministic cross sector is not written as a matched force coupling between ∇ rµ and ∂ τ µ, but rather through the density-curvature proxy ˆ∇ 2ρ i: vcross τ ,i = gχ Mrτ (ρ i) kBT ˆ∇ 2ρ i, (A3) vcross r,i = g χ Mτ r...
work page 2000
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.