Recognition: 2 theorem links
· Lean TheoremNeuromodulation supports robust rhythmic pattern transitions in degenerate central pattern generators with fixed connectivity
Pith reviewed 2026-05-10 17:45 UTC · model grok-4.3
The pith
An adaptive neuromodulation controller in low-dimensional feedback gain space robustly enforces gait transitions in degenerate central pattern generators despite fixed connectivity and large parametric variability.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
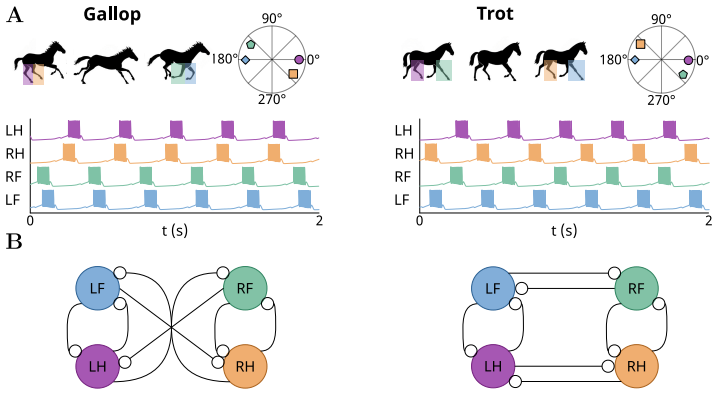
Equivariant bifurcation theory supplies necessary symmetry conditions on neuromodulatory projection topology that permit target gaits. An adaptive neuromodulation controller working in a low-dimensional feedback gain space then enforces gait transitions robustly in conductance-based neuron models even under large parametric variability, as shown by reliable gallop-to-trot transitions in a quadrupedal gait-control problem across 200 degenerate networks with up to fivefold conductance variability.
What carries the argument
Adaptive neuromodulation controller operating in low-dimensional feedback gain space, whose action is constrained by symmetry conditions on neuromodulatory projection topology obtained from equivariant bifurcation theory.
If this is right
- Reliable gallop-to-trot transitions become achievable in quadrupedal central pattern generator models with fixed connectivity.
- The controller maintains performance across 200 distinct networks that differ by up to fivefold in conductance parameters.
- Rapid rhythmic reconfiguration occurs without any modification to the underlying network connections.
- The same low-dimensional adaptation mechanism works for conductance-based neuron models exhibiting structured degeneracy.
Where Pith is reading between the lines
- The same symmetry-guided adaptation may apply to other rhythmic biological systems, such as respiratory central pattern generators, where fast transitions are also required.
- Designers of robotic locomotion controllers could adopt the low-dimensional gain-space approach to achieve robustness against component variability.
- Networks whose neuromodulatory projections do not satisfy the symmetry conditions should show systematic failure of gait transitions under this controller.
- The method could be extended to networks with more complex or higher-dimensional connectomes to test how broadly the bifurcation-derived symmetry conditions scale.
Load-bearing premise
The symmetry conditions on neuromodulatory projection topology derived from equivariant bifurcation theory must hold in order to enable the target gaits amid structured neuronal degeneracy.
What would settle it
A simulation or experiment in which the adaptive controller is applied to a network whose neuromodulatory projection topology violates the derived symmetry conditions, after which the controller fails to produce the target gait transitions despite gain adaptation.
Figures





read the original abstract
Many essential biological functions, such as breathing and locomotion, rely on the coordination of robust and adaptable rhythmic patterns, governed by specific network architectures known as connectomes. Rhythmic adaptation is often linked to slow structural modifications of the connectome through synaptic plasticity, but such mechanisms are too slow to support rapid, localized rhythmic transitions. Here, we propose a neuromodulation-based control architecture for dynamically reconfiguring rhythmic activity in networks with fixed connectivity. The key control challenge is to achieve reliable rhythm switching despite neuronal degeneracy, a form of structured variability where widely different parameter combinations produce similar functional output. Using equivariant bifurcation theory, we derive necessary symmetry conditions on the neuromodulatory projection topology for the existence of target gaits. We then show that an adaptive neuromodulation controller, operating in a low-dimensional feedback gain space, robustly enforces gait transitions in conductance-based neuron models despite large parametric variability. The framework is validated in simulation on a quadrupedal gait control problem, demonstrating reliable gallop-to-trot transitions across 200 degenerate networks with up to fivefold conductance variability.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that symmetry conditions on neuromodulatory projection topology, derived via equivariant bifurcation theory, enable an adaptive low-dimensional feedback controller to robustly drive gait transitions (such as gallop-to-trot) in fixed-connectivity conductance-based CPG networks, even when neuronal parameters exhibit degeneracy with up to fivefold conductance variability across 200 simulated networks.
Significance. If the central claim holds, the work would advance the application of equivariant dynamical systems to biological motor control by showing how neuromodulation can achieve rapid, reliable pattern switching without altering connectivity or relying on slow plasticity. The combination of bifurcation-theoretic derivation with extensive simulation validation across degenerate parameter sets is a notable strength, offering both mechanistic insight and a practical control architecture for degenerate systems.
major comments (2)
- [Section 3] Section 3 (equivariant bifurcation analysis): The necessary symmetry conditions on the neuromodulatory projection are derived under the assumption that the vector field is equivariant under the connectome symmetry group. However, independent up to fivefold variations in maximal conductances across neurons (as used to generate the 200 degenerate networks) generally break this equivariance unless the variability is itself group-invariant. The manuscript does not report an explicit verification that equivariance (or the target bifurcations) is preserved for each sampled parameter set.
- [Section 5] Section 5 (numerical validation): While the simulations demonstrate successful transitions in all 200 networks, the paper provides no details on how the degenerate parameter sets were sampled to ensure they remain functionally equivalent yet test the symmetry conditions, nor any post-simulation check that the adaptive controller's performance relies on the theoretically derived symmetries rather than incidental numerical effects.
minor comments (2)
- [Abstract] The abstract and introduction could more precisely define the range and statistical distribution of the conductance variability (e.g., uniform vs. log-normal sampling) to aid reproducibility.
- [Section 2] Notation for the feedback gain space and neuromodulatory projection matrix would benefit from an early summary table or diagram to clarify the low-dimensional controller structure.
Simulated Author's Rebuttal
We thank the referee for the careful and constructive review. The comments identify important points regarding the preservation of symmetry under parameter degeneracy and the need for greater transparency in the numerical methods. We will revise the manuscript to address these by adding explicit verifications and methodological details. Our point-by-point responses follow.
read point-by-point responses
-
Referee: [Section 3] Section 3 (equivariant bifurcation analysis): The necessary symmetry conditions on the neuromodulatory projection are derived under the assumption that the vector field is equivariant under the connectome symmetry group. However, independent up to fivefold variations in maximal conductances across neurons (as used to generate the 200 degenerate networks) generally break this equivariance unless the variability is itself group-invariant. The manuscript does not report an explicit verification that equivariance (or the target bifurcations) is preserved for each sampled parameter set.
Authors: We agree that independent parameter variations can perturb exact equivariance of the vector field. The bifurcation analysis derives necessary conditions assuming the nominal symmetric connectome, which then inform the topology of the neuromodulatory projections. The adaptive controller is subsequently shown to drive transitions in simulation. To address the concern directly, we will add to the revised manuscript an explicit post-sampling verification: for each of the 200 networks we will quantify the deviation from group equivariance (via the norm of the commutator between the vector field and the group action) and confirm that the target bifurcations remain accessible under the controller. We will also clarify the sampling constraints that preserve functional rhythmic output. revision: yes
-
Referee: [Section 5] Section 5 (numerical validation): While the simulations demonstrate successful transitions in all 200 networks, the paper provides no details on how the degenerate parameter sets were sampled to ensure they remain functionally equivalent yet test the symmetry conditions, nor any post-simulation check that the adaptive controller's performance relies on the theoretically derived symmetries rather than incidental numerical effects.
Authors: We accept that additional methodological detail is required. In the revised Section 5 we will describe the sampling procedure used to generate the 200 networks, including the ranges and constraints applied to ensure each set produces stable rhythmic activity without modulation (functional equivalence). We will also add post-simulation analyses, such as ablation of the symmetry in the projection topology and comparison of transition success rates, to demonstrate that performance depends on the equivariant conditions rather than incidental factors. revision: yes
Circularity Check
No significant circularity; derivation relies on external theory and simulation validation
full rationale
The paper derives necessary symmetry conditions from equivariant bifurcation theory (an established external framework) and validates the adaptive controller via direct simulation across 200 degenerate networks. No load-bearing step reduces a claim to a self-definition, a fitted parameter renamed as prediction, or a self-citation chain that is unverified. The central result combines theoretical derivation with numerical evidence on fixed-connectivity models, remaining self-contained against the stated assumptions without tautological reduction.
Axiom & Free-Parameter Ledger
free parameters (1)
- feedback gains
axioms (1)
- domain assumption Equivariant bifurcation theory provides necessary symmetry conditions on neuromodulatory projection topology for target gaits
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Using equivariant bifurcation theory, we derive necessary symmetry conditions on the neuromodulatory projection topology for the existence of target gaits... Theorem 1 (Theorem 3.4 in [26])... K is an isotropy subgroup; dim Fix(K) >=2
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
neuronal degeneracy... widely different parameter combinations produce similar functional output... up to fivefold conductance variability
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Neural mechanisms underlying the generation of the lobster gastric mill motor pattern
A. Selverston, A. Sz ¨ucs, R. Huerta, R. D. Pinto and M. B. Reyes, “Neural mechanisms underlying the generation of the lobster gastric mill motor pattern”,Front. Neural Circuits, vol. 3, p. 608, 2009
2009
-
[2]
Central pattern generators and the control of rhythmic movements
E. Marder and D. Bucher, “Central pattern generators and the control of rhythmic movements”,Curr. Biol., vol. 11, no. 23, pp. R986–R996, 2001
2001
-
[3]
Programmable coupled oscillators for synchronized locomotion
S. Dutta et al., “Programmable coupled oscillators for synchronized locomotion”,Nat. Commun., vol. 10, no. 1, p. 3299, 2019
2019
-
[4]
Principles of rhythmic motor pattern generation
E. Marder and R. L. Calabrese, “Principles of rhythmic motor pattern generation”,Physiol. Rev., vol. 76, no. 3, pp. 687–717, 1996
1996
-
[5]
From the connectome to brain function
C. I. Bargmann and E. Marder, “From the connectome to brain function”,Nat. Methods, vol. 10, no. 6, pp. 483–490, 2013
2013
-
[6]
Hardware implementation of a CPG-based locomotion control for quadruped robots
J. H. Barron-Zambrano, C. Torres-Huitzil and B. Girau, “Hardware implementation of a CPG-based locomotion control for quadruped robots”, inProc. Int. Conf. Artificial Neural Networks, Thessaloniki, Greece, pp. 276–285, 2010
2010
-
[7]
Modeling of a neural pattern generator with coupled nonlinear oscillators
J. Bay and H. Hemami, “Modeling of a neural pattern generator with coupled nonlinear oscillators”,IEEE Trans. Biomed. Eng., vol. BME- 34, no. 4, pp. 297–306, 2007
2007
-
[8]
A survey on CPG-inspired control models and system implementation
J. Yu, M. Tan, J. Chen and J. Zhang, “A survey on CPG-inspired control models and system implementation”,IEEE Trans. Neural Netw. Learn. Syst., vol. 25, no. 3, pp. 441–456, 2013
2013
-
[9]
Phenomenological models of synaptic plasticity based on spike timing
A. Morrison, M. Diesmann and W. Gerstner, “Phenomenological models of synaptic plasticity based on spike timing”,Biol. Cybern., vol. 98, pp. 459–478, 2008
2008
-
[10]
Ion channel degeneracy, variability, and covariation in neuron and circuit resilience
J. M. Goaillard and E. Marder, “Ion channel degeneracy, variability, and covariation in neuron and circuit resilience”,Annu. Rev. Neurosci., vol. 44, no. 1, pp. 335–357, 2021
2021
-
[11]
Degeneracy rules!
A. A. Prinz, “Degeneracy rules!”,J. Physiol., vol. 595, no. 8, p. 2409, 2017
2017
-
[12]
Designing reliable systems from unreliable components: The challenges of transistor variability and degradation
S. Borkar, “Designing reliable systems from unreliable components: The challenges of transistor variability and degradation”,IEEE Micro, vol. 25, no. 6, pp. 10–16, 2006
2006
-
[13]
Neuromodulation of circuits with variable parameters: Single neurons and small circuits reveal principles of state-dependent and robust neuromodulation
E. Marder, T. O’Leary and S. Shruti, “Neuromodulation of circuits with variable parameters: Single neurons and small circuits reveal principles of state-dependent and robust neuromodulation”,Annu. Rev. Neurosci., vol. 37, no. 1, pp. 329–346, 2014
2014
-
[14]
Dimensionality reduction of neuronal degeneracy reveals two interfering physiological mecha- nisms
A. Fyon, A. Franci, P. Sacr ´e and G. Drion, “Dimensionality reduction of neuronal degeneracy reveals two interfering physiological mecha- nisms”,PNAS Nexus, vol. 3, no. 10, p. pgae415, 2024
2024
-
[15]
A. Fyon and G. Drion, “Neuromodulation and homeostasis: comple- mentary mechanisms for robust neural function”, arXiv:2412.04172, 2024
work page internal anchor Pith review Pith/arXiv arXiv 2024
-
[16]
Fast reconstruction of degenerate populations of conductance-based neuron models from spike times
J. Brandoit, D. Ernst, G. Drion and A. Fyon, “Fast reconstruction of degenerate populations of conductance-based neuron models from spike times”, arXiv:2509.12783, 2025
-
[17]
Central pattern generators for locomotion control in animals and robots: A review
A. J. Ijspeert, “Central pattern generators for locomotion control in animals and robots: A review”,Neural Netw., vol. 21, no. 4, pp. 642– 653, 2008
2008
-
[18]
Circuit robustness to temperature perturbation is altered by neuromodulators
S. A. Haddad and E. Marder, “Circuit robustness to temperature perturbation is altered by neuromodulators”,Neuron, vol. 100, no. 3, pp. 609–623, 2018
2018
-
[19]
Central pattern generators
D. Bucher, G. Haspel, J. Golowasch and F. Nadim, “Central pattern generators”,eLS, vol. 11, pp. 1–12, 2015
2015
-
[20]
The components of membrane conductance in the giant axon of Loligo
A. L. Hodgkin and A. F. Huxley, “The components of membrane conductance in the giant axon of Loligo”,J. Physiol., vol. 116, no. 4, p. 473, 1952
1952
-
[21]
Modeling the mammalian locomotor CPG: insights from mistakes and perturbations
D. A. McCrea and I. A. Rybak, “Modeling the mammalian locomotor CPG: insights from mistakes and perturbations”,Prog. Brain Res., vol. 165, pp. 235–253, 2007
2007
-
[22]
Neuromorphic silicon neuron circuits
G. Indiveri et al., “Neuromorphic silicon neuron circuits”,Front. Neurosci., vol. 5, p. 73, 2011
2011
-
[23]
Ultra-low power silicon neuron circuit for extreme-edge neuromorphic intelligence
A. Rubino, M. Payvand and G. Indiveri, “Ultra-low power silicon neuron circuit for extreme-edge neuromorphic intelligence”, inProc. 26th IEEE Int. Conf. Electronics, Circuits and Systems (ICECS), Genoa, Italy, pp. 458–461, 2019
2019
-
[24]
Advancements in trajectory optimization and model predictive control for legged systems
E. Mingo Hoffman, C. Zhou and M. Parigi Polverini, “Advancements in trajectory optimization and model predictive control for legged systems”,Front. Robot. AI, vol. 9, p. 1002552, 2022
2022
-
[25]
Next generation legged robot locomotion: A review on control techniques
S. S. Kotha et al., “Next generation legged robot locomotion: A review on control techniques”,Heliyon, vol. 10, no. 18, p. e37237, 2024
2024
-
[26]
Golubitsky and I
M. Golubitsky and I. Stewart,The Symmetry Perspective: From Equilibrium to Chaos in Phase Space and Physical Space. Basel, Switzerland: Birk ¨auser, 2003
2003
-
[27]
A modular network for legged locomotion
M. Golubitsky, I. Stewart, P.-L. Buono and J. J. Collins, “A modular network for legged locomotion”,Physica D, vol. 115, no. 1–2, pp. 56–72, 1998
1998
-
[28]
Coupled nonlinear oscillators and the symmetries of animal gaits
J. J. Collins and I. Stewart, “Coupled nonlinear oscillators and the symmetries of animal gaits”,J. Nonlinear Sci., vol. 3, no. 1, pp. 349– 392, 1993
1993
-
[29]
The neuromuscular junction
A. G. Engel, “The neuromuscular junction”,Handb. Clin. Neurol., vol. 91, pp. 103–148, 2008
2008
-
[30]
Reliable neuromodulation from adaptive control of ion channel expression
A. Fyon, P. Sacr ´e, A. Franci and G. Drion, “Reliable neuromodulation from adaptive control of ion channel expression”,IFAC-PapersOnLine, vol. 56, no. 2, pp. 458–463, 2023
2023
-
[31]
Half-center oscillators underlying rhythmic move- ments
R. L. Calabrese, “Half-center oscillators underlying rhythmic move- ments”,Nature, vol. 261, pp. 146–148, 1995
1995
-
[32]
Spiking control systems
R. Sepulchre, “Spiking control systems”,Proc. IEEE, vol. 110, no. 5, pp. 577–589, 2022
2022
-
[33]
Cell types, network homeostasis, and pathological compensation from a biologi- cally plausible ion channel expression model
T. O’Leary, A. H. Williams, A. Franci and E. Marder, “Cell types, network homeostasis, and pathological compensation from a biologi- cally plausible ion channel expression model”,Neuron, vol. 82, no. 4, pp. 809–821, 2014
2014
-
[34]
A model neuron with activity-dependent conductances regulated by multiple calcium sensors
Z. Liu, J. Golowasch, E. Marder and L. F. Abbott, “A model neuron with activity-dependent conductances regulated by multiple calcium sensors”,J. Neurosci., vol. 18, no. 7, pp. 2309–2320, 1998
1998
-
[35]
Central pattern generating neurons simultaneously express fast and slow rhythmic activities in the stomatogastric ganglion
D. Bucher, A. L. Taylor and E. Marder, “Central pattern generating neurons simultaneously express fast and slow rhythmic activities in the stomatogastric ganglion”,J. Neurophysiol., vol. 95, no. 6, pp. 3617– 3632, 2006
2006
-
[36]
Cellular switches orchestrate rhythmic circuits
G. Drion, A. Franci and R. Sepulchre, “Cellular switches orchestrate rhythmic circuits”,Biol. Cybern., vol. 113, no. 1, pp. 71–82, 2019
2019
-
[37]
Neural control of brain state
E. Zagha and D. A. McCormick, “Neural control of brain state”,Curr. Opin. Neurobiol., vol. 29, pp. 178–186, 2014
2014
-
[38]
Dynamic input conductances shape neuronal spiking
G. Drion, A. Franci, J. Dethier and R. Sepulchre, “Dynamic input conductances shape neuronal spiking”,eNeuro, vol. 2, no. 1, 2015
2015
-
[39]
A review on model reference adaptive control of robotic manipulators
D. Zhang and B. Wei, “A review on model reference adaptive control of robotic manipulators”,Annu. Rev. Control, vol. 43, pp. 188–198, 2017
2017
-
[40]
Ion channel degeneracy enables robust and tunable neuronal firing rates
G. Drion, T. O’Leary and E. Marder, “Ion channel degeneracy enables robust and tunable neuronal firing rates”,PNAS, vol. 112, no. 38, pp. E5361–E5370, 2015
2015
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.