Kinetics of segregation of topologically-modified ring polymers in cylindrical confinement
Pith reviewed 2026-05-07 14:15 UTC · model grok-4.3
The pith
Adding more internal loops to ring polymers shortens their segregation time under cylindrical confinement by increasing entropic repulsion.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
With certain caveats, increasing the number of loops in topologically modified ring polymers decreases the time required for segregation under cylindrical confinement. This occurs because the loops raise the entropic repulsion between the polymers. The contour length of the loops and the mutual orientation of the anisotropic chains in the initial configurations also determine the segregation time.
What carries the argument
Entropic repulsion between two topologically modified ring polymers, enhanced by internal loops created through monomer crosslinking, under cylindrical confinement.
If this is right
- Segregation completes faster as the number of loops rises.
- Longer contour lengths of the loops alter the segregation kinetics.
- The initial mutual orientation of the anisotropic chains influences how quickly segregation occurs.
- The added loops increase the effective repulsion that drives the two rings apart.
Where Pith is reading between the lines
- The placement of loops near the origin of replication in real E. coli chromosomes may be tuned to promote faster segregation.
- Topological loop modifications could be engineered in synthetic confined polymers to control separation rates.
- The results point to a general role for internal loops in speeding polymer separation inside narrow channels.
- Varying the cylinder radius in follow-up simulations would test how confinement strength interacts with the loop effect.
Load-bearing premise
The bead-spring model with crosslinking at selected locations and the chosen initial configurations accurately represent the entropic repulsion and segregation dynamics of real topologically modified chromosomal DNA in E. coli under cylindrical confinement.
What would settle it
An observation that segregation time does not decrease, or that measured repulsion does not increase, when the number of loops is raised in either alternative polymer models or experimental confined-ring systems would falsify the central relationship.
Figures












read the original abstract
In Escherichia coli (E. coli), entropic repulsion between the two daughter DNA ring polymers under cylindrical confinement is believed to be an important factor governing chromosomal segregation. The repulsion can be enhanced by topological modifications, i.e., by the introduction of internal loops at certain locations along the contour of the circular DNA. However, the effect of topological modifications on the rate of segregation of ring polymers remains unclear. Therefore, we systematically varied the number and the contour length of loops introduced at selected locations by crosslinking monomers. The appropriate crosslinking was motivated by observations that extruded loops are located mainly near the origin of replication (ori-proximal) region of the E. coli chromosome. This resulted in the chains becoming intrinsically anisotropic. Using Langevin dynamics simulations of these topologically modified bead-spring polymers, we calculated the time required for segregation under cylinder confinement. With certain caveats, we found that increasing the number of loops resulted in a decrease in the time of segregation. In line with past work, we propose that this is due to the increase in the entropic repulsion between the polymers upon increasing the number of loops. In addition to the number of loops, the contour length of the loops and the mutual orientation of the (anisotropic) chains in the initial configurations played a role in determining the time of segregation.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses Langevin dynamics simulations of bead-spring ring polymers under cylindrical confinement to study how introducing internal loops (via monomer crosslinking at selected ori-proximal locations) affects segregation kinetics. The central claim is that, with certain caveats, increasing the number of loops decreases the time to segregation, which the authors attribute to enhanced entropic repulsion between the chains; loop contour length and the mutual orientation of the resulting anisotropic chains are also reported to influence segregation times.
Significance. If the reported trend with loop number is robust to controls for initial orientation, the work would strengthen the entropic-repulsion mechanism for bacterial chromosome segregation by showing that specific topological modifications can accelerate the process. The systematic variation of loop number and length, together with the explicit motivation from E. coli ori-proximal extrusion, provides a concrete link between polymer topology and confinement-driven dynamics that can be tested against future experiments or more detailed models.
major comments (2)
- [initial configurations] § on initial configurations and anisotropy: The abstract states that 'the mutual orientation of the (anisotropic) chains in the initial configurations played a role in determining the time of segregation.' Because loop placement itself renders the chains anisotropic, different loop numbers inherently change the possible orientations. The manuscript must demonstrate that orientations were either fixed across loop numbers or systematically sampled and averaged; otherwise the observed decrease in segregation time cannot be cleanly attributed to increased entropic repulsion rather than orientation-dependent packing or collision statistics.
- [results] Results section on segregation times: The phrase 'with certain caveats' is used for the trend of decreasing segregation time with loop number, yet the manuscript does not quantify how these caveats (including orientation effects) alter the statistical significance or magnitude of the trend. Error bars, number of independent runs, and a direct comparison of segregation-time distributions for matched versus unmatched orientations are needed to establish that the central claim is load-bearing.
minor comments (2)
- [abstract] The abstract would be clearer if it stated the specific range of loop numbers and contour lengths examined.
- [figures] Figure captions should explicitly note whether error bars represent standard error over independent trajectories or another measure.
Simulated Author's Rebuttal
We thank the referee for their careful reading of our manuscript and for highlighting important points regarding initial configurations and the quantification of caveats. We have revised the manuscript to provide additional details on simulation protocols, include statistical measures, and present new control analyses that address the concerns while preserving the central findings on entropic repulsion.
read point-by-point responses
-
Referee: [initial configurations] § on initial configurations and anisotropy: The abstract states that 'the mutual orientation of the (anisotropic) chains in the initial configurations played a role in determining the time of segregation.' Because loop placement itself renders the chains anisotropic, different loop numbers inherently change the possible orientations. The manuscript must demonstrate that orientations were either fixed across loop numbers or systematically sampled and averaged; otherwise the observed decrease in segregation time cannot be cleanly attributed to increased entropic repulsion rather than orientation-dependent packing or collision statistics.
Authors: We agree that a clear demonstration is required to separate orientation effects from the topological contribution. In the original simulations, initial configurations were prepared by inserting the polymers into the cylinder with randomized but bounded orientations (specifically, the angle between the principal axes of the two chains was sampled uniformly over [0, π/2] for each topology). To address the referee's point directly, we have added a dedicated subsection in the Methods describing the generation protocol and performed a new control study in which orientations were fixed to identical values across all loop numbers. The revised Results section now reports that the decrease in mean segregation time with increasing loop number remains statistically significant (p < 0.01) under fixed-orientation conditions, supporting attribution to enhanced entropic repulsion. A supplementary figure shows the full distribution of segregation times for matched versus unmatched orientations. revision: yes
-
Referee: [results] Results section on segregation times: The phrase 'with certain caveats' is used for the trend of decreasing segregation time with loop number, yet the manuscript does not quantify how these caveats (including orientation effects) alter the statistical significance or magnitude of the trend. Error bars, number of independent runs, and a direct comparison of segregation-time distributions for matched versus unmatched orientations are needed to establish that the central claim is load-bearing.
Authors: We accept that the original presentation of caveats was insufficiently quantitative. The revised manuscript now states that each data point is the average of 50 independent Langevin trajectories, with error bars indicating the standard error of the mean. We have replaced the phrase 'with certain caveats' with a quantified statement: the observed reduction in segregation time with loop number is robust (approximately 25–40 % decrease from 0 to 4 loops) except for the longest loops (contour length > 20 % of the ring) and for perfectly anti-aligned initial orientations, where the effect is attenuated but still present. A new panel in Figure 3 directly compares the cumulative distribution functions of segregation times for matched-orientation versus random-orientation ensembles, confirming that orientation modulates absolute times but does not reverse the trend with loop number. These additions establish that the central claim remains load-bearing after accounting for the identified caveats. revision: yes
Circularity Check
No circularity: segregation times obtained directly from Langevin dynamics simulations
full rationale
The paper's central result is a numerical trend in segregation times extracted from explicit Langevin dynamics trajectories of bead-spring ring polymers with varying numbers of crosslinked loops. No equations are presented whose outputs are algebraically identical to their inputs, no parameters are fitted to a subset of data and then relabeled as predictions, and no load-bearing premise rests on a self-citation whose content is itself unverified within the work. The attribution of the trend to enhanced entropic repulsion is offered as an interpretation aligned with prior literature rather than a deductive step that presupposes the observed outcome. The acknowledgment that initial mutual orientations affect segregation times is stated explicitly and does not create a self-referential loop. The derivation chain therefore remains self-contained against external simulation benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Langevin dynamics with a bead-spring model sufficiently captures the entropic and topological effects governing segregation of ring polymers in cylindrical confinement.
Reference graph
Works this paper leans on
-
[1]
The polymers were further equilibrated until t = t0 while subject to the ad- ditional constraints imposed during the four initializa- tion protocols (refer Fig. 2 and see below for additional details). The time t = t0 marked the end of the ini- tialization run after which the constraints were removed and the polymers allowed to segregate. For each of the ...
-
[2]
In other words, monomer i of one polymer interacted with monomer i of the other polymer for all 1 ≤ i ≤ N (see Fig. 2(c)). Note however that κ = 10 ǫ/σ 2 and r0 = 3 σ was used. These values allowed spatial fluctuations of the monomers but maintained suf- ficient overlap between the two polymers. (iv) Mutual-Attraction: As before, monomers belong- ing to the...
-
[3]
In this “steady state”, the polymers occupied different halves, with their COMs located approximately near L/ 4 and 3 L/ 4 along the axis. Occasional decreases in DCOM (gray arrows in Fig
-
[4]
corresponded to tran- sient remixing. Ideally, one would like to define the time of segregation as the time beyond which the two polymers 6 remained segregated indefinitely. In microscopic systems, such as the ones under consideration, remixing is possible due to thermal fluctuations given sufficient time. There- fore, in practice, to define the time of segrega...
-
[5]
The time at which the aforementioned condi- tion was satisfied for the i-th time was denoted as T i F . (ii) Starting from each T i F , ⟨DCOM⟩, the time-average over a time duration Ts = 5N 2τ0, must satisfy ⟨DCOM⟩ > dS. Here dS is another threshold. In the rare instances where 10 N 2τ0 − T n F < 5N 2τ0, the time average was cal- culated for Ts = 10 N 2τ0 ...
-
[6]
For each run, the time of segregation, τseg, was determined using the criterion discussed in Section 2 E. Varying loop lengths in Arc-1-1: N = 200 Before examining the different topologies, we investi- gated the effect of varying the number of monomers in the small loop, NS, on τseg of N = 200 Arc-1-1 polymers. For example, a polymer with NS = 60, denoted b...
-
[7]
The whiskers extended from the edges of the box to the data points lying within 1
Mean (blue diamonds), second quartile or the median (Q 2, blue line within the box), third quar- tile (Q 3, top edge of the box), first quartile (Q 1, bot- tom edge), and the number of outliers are shown. The whiskers extended from the edges of the box to the data points lying within 1 . 5 × IQR, where IQR ≡ Q3 − Q1 is the interquartile range. Data points ...
- [8]
-
[9]
For the Glued-Monomers and Ladder- like protocols, τseg decreased with increasing number of small loops, consistent with that observed for N = 200. However, τseg obtained when the initial configurations were generated using the Recenter-COM and the Mutual- Attraction protocols deviated from this trend, with Arc- 1-5 and Arc-1-10 exhibiting unexpectedly lar...
-
[10]
Examin- ing the COMs of the cluster of small loops and the large loop of each polymer individually enabled us to discern the behavior of the individual sections and identify over- laps during the different stages of segregation. The char- acteristics of the motion of the clusters of small loops (or occasionally the large loops) toward the poles of the cyli...
-
[11]
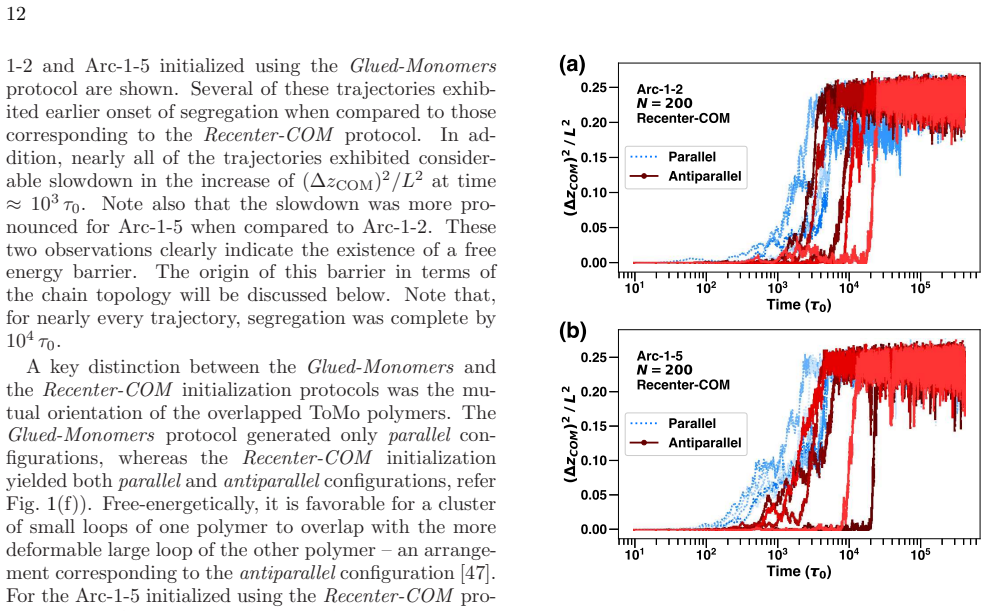
Similar data for L/D = 5 was provided in Fig. 9. For all of the topologies, (∆ zCOM )2 appeared to increase with time while being subject to significant fluctuations. For time ⩽ 102τ0, ∆ z2 COM ⩽ σ 2, indicating that the polymers ex- hibited substantial overlap. The increase in (∆ zCOM )2 with time indicated the progress of segregation. Nearly all of Arc-0 ...
-
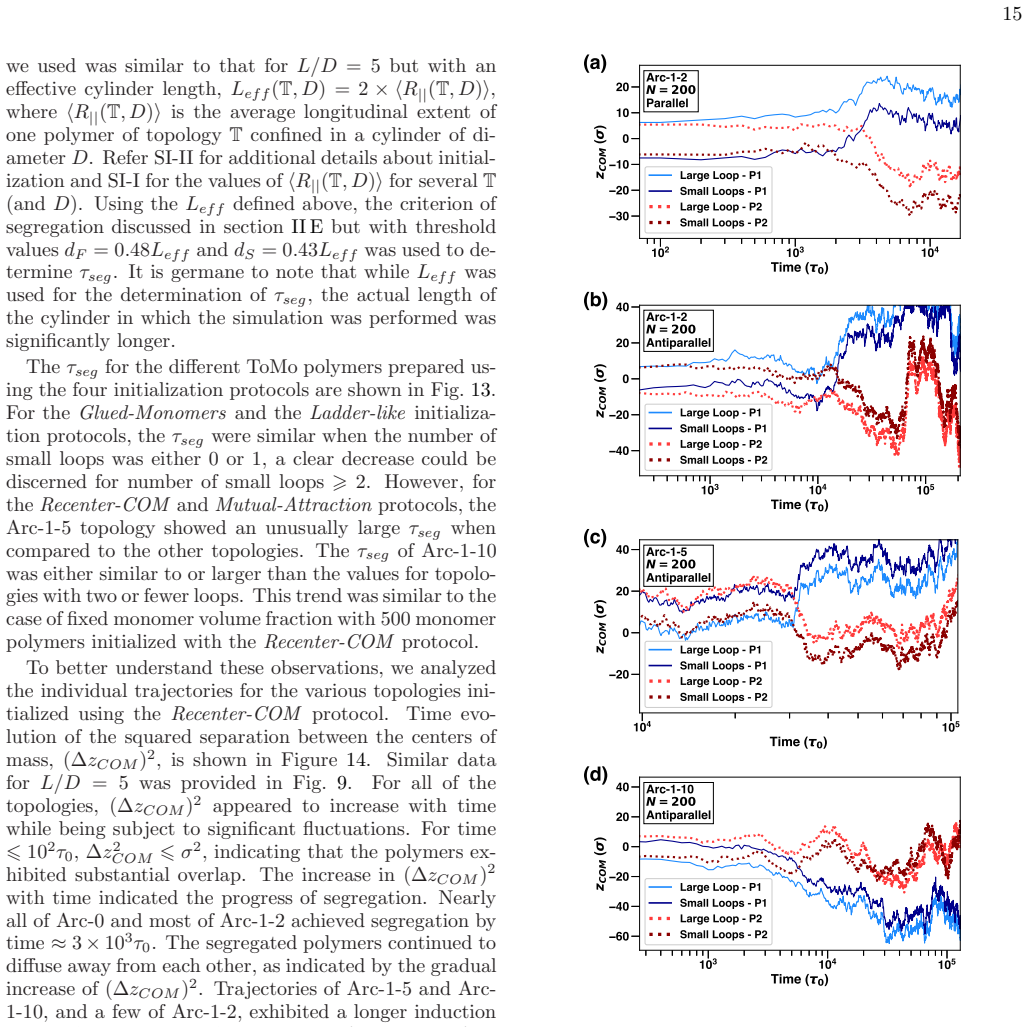
[12]
can be attributed to the antipar- 10 2 10 3 10 4 Time ( τ 0 ) −30 −20 −10 0 10 20 z COM ( σ ) Arc -1-2 N = 200 Parallel Large Loop - P1 Small Loops - P1 Large Loop - P2 Small Loops - P2 (a) 10 3 10 4 10 5 Time ( τ 0 ) −40 −20 0 20 40 z COM ( σ ) Arc -1-2 N = 200 Antiparallel Large Loop - P1 Small Loops - P1 Large Loop - P2 Small Loops - P2 (b) 10 4 10 5 T...
-
[13]
In Fig. 15 (a), corresponding to the initial configuration of two Arc-1- 2 polymers being parallel, the segregation occurred by the displacement of polymer P1 such that its large loop was ahead of the small loops while polymer P2 moved in the opposite direction with the small loops ahead of the large loop. Unless a parallel to antiparallel flip occurred at ...
work page 1973
- [14]
- [15]
- [16]
- [17]
-
[18]
S. Jun, A. Arnold, and B.-Y. Ha, Physical Review Letters 98, 128303 (2007)
work page 2007
- [19]
-
[20]
Y. Du, H. Jiang, and Z. Hou, The Journal of Chemical Physics 149, 244906 (2018)
work page 2018
- [21]
- [22]
-
[23]
Y. Jung, C. Jeon, J. Kim, H. Jeong, S. Jun, and B.-Y. Ha, Soft Matter 8, 2095 (2012)
work page 2095
-
[24]
J. Shin, A. G. Cherstvy, and R. Metzler, New Journal of Physics 16, 053047 (2014)
work page 2014
- [25]
-
[26]
J. M. Polson and L. G. Montgomery, The Journal of Chemical Physics 141, 164902 (2014)
work page 2014
-
[27]
J. M. Polson and D. R.-M. Kerry, Soft Matter 14, 6360 (2018)
work page 2018
-
[28]
J. Pelletier, K. Halvorsen, B.-Y. Ha, R. Paparcone, S. J. Sandler, C. L. Woldringh, W. P. Wong, and S. Jun, Proceedings of the National Academy of Sciences 109, E2649 (2012)
work page 2012
- [29]
- [30]
-
[31]
X. Wang, X. Liu, C. Possoz, and D. J. Sherratt, Genes & Development 20, 1727 (2006)
work page 2006
-
[32]
J. Cass, N. Kuwada, B. Traxler, and P. Wiggins, Biophysical Journal 110, 2597 (2016)
work page 2016
-
[33]
A. Kuzminov, Molecular Microbiology 90, 214 (2013) , https://onlinelibrary.wiley.com/doi/pdf/10.1111/mmi.12372
-
[34]
A. Badrinarayanan, T. B. Le, and M. T. Laub, Annual Review of Cell and Developmental Biology 31, 171 (2015)
work page 2015
-
[35]
C. L. Woldringh, Life 14, 660 (2024)
work page 2024
-
[36]
N. Kleckner, J. K. Fisher, M. Stouf, M. A. White, D. Bates, and G. Witz, 18 Current Opinion in Microbiology 22, 127 (2014)
work page 2014
-
[37]
R. T. Dame, F.-Z. M. Rashid, and D. C. Grainger, Nature Reviews Genetics 21, 227 (2019)
work page 2019
- [38]
-
[39]
V. S. Lioy, I. Junier, and F. Boccard, Annual review of microbiology 75, 541 (2021)
work page 2021
- [40]
- [41]
-
[42]
J. M¨ akel¨ a, S. Uphoff, and D. J. Sherratt, Proceedings of the National Academy of Sciences 118, e2022078118 (2021)
work page 2021
- [43]
-
[44]
J. Harju and C. P. Broedersz, Molecular Microbiology (2024), 10.1111/mmi.15257
-
[45]
S. Brahmachari and J. F. Marko, Proceedings of the National Academy of Sciences 116, 24956 (2019) , https://www.pnas.org/content/116/50/24956.full.pdf
work page 2019
- [46]
- [47]
- [48]
-
[49]
V. S. Lioy, A. Cournac, M. Marbouty, S. Duigou, J. Mozziconacci, O. Esp´ eli, F. Boccard, and R. Koszul, Cell 172, 771 (2018)
work page 2018
-
[50]
T. Gaal, B. P. Bratton, P. Sanchez-Vazquez, A. Sliwicki, K. Sliwicki, A. Vegel, R. Pannu, and R. L. Gourse, Genes & Development 30, 2272 (2016)
work page 2016
-
[51]
D. D’Asaro, M. M. Tortora, C. Vaillant, J.-M. Arbona, and D. Jost, Physical Review X 14, 041020 (2024)
work page 2024
-
[52]
M. Di Stefano, J. Paulsen, D. Jost, and M. A. Marti-Renom, Current Opinion in Genetics & Development 67, 25–32 (2021)
work page 2021
- [53]
-
[54]
C. A. Brackley, S. Taylor, A. Papanto- nis, P. R. Cook, and D. Marenduzzo, Proceedings of the National Academy of Sciences 110, E3605 (2013)
work page 2013
-
[55]
C. A. Brackley, J. Johnson, S. Kelly, P. R. Cook, and D. Marenduzzo, Nucleic Acids Research 44, 3503–3512 (2016)
work page 2016
-
[56]
A. Maji, J. A. Ahmed, S. Roy, B. Chakrabarti, and M. K. Mitra, Biophysical Journal 118, 3041 (2020)
work page 2020
- [57]
-
[58]
J. F. Marko, Physical Review E 79, 051905 (2009)
work page 2009
- [59]
-
[60]
S. Bhandarkar, D. Mitra, J. Horbach, and A. Chatterji, Physical Review E 113, 045401 (2026)
work page 2026
-
[61]
S. Brahmachari, A. B. Oliveira, M. F. Mello, V. G. Contessoto, and J. N. Onuchic, Proceedings of the National Academy of Sciences 123, e2535321123
- [62]
-
[63]
K. Roychoudhury, S. Pande, I. S. Shashank, D. Mitra, and A. Chatterji, Physical Review E 112, 055409 (2025)
work page 2025
-
[64]
M. P. Allen and D. J. Tildesley, Computer simulation of liquids , second edition ed. (Oxford University Press, Oxford, 2017)
work page 2017
-
[65]
K. Kremer and G. S. Grest, The Journal of Chemical Physics 92, 5057 (1990)
work page 1990
-
[66]
J. D. Weeks, D. Chandler, and H. C. Andersen, The Journal of Chemical Physics 54, 5237 (1971)
work page 1971
-
[67]
A. P. Thompson, H. M. Aktulga, R. Berger, D. S. Bolin- tineanu, W. M. Brown, P. S. Crozier, P. J. in ’t Veld, A. Kohlmeyer, S. G. Moore, T. D. Nguyen, R. Shan, M. J. Stevens, J. Tranchida, C. Trott, and S. J. Plimp- ton, Comp. Phys. Comm. 271, 108171 (2022)
work page 2022
-
[68]
M. A. Carignano, M. Kroeger, L. M. Almassalha, V. Agrawal, W. S. Li, E. M. Pujadas-Liwag, R. J. Nap, V. Backman, and I. Szleifer, eLife 13, RP97604 (2024)
work page 2024
-
[69]
H. Doshi and A. Chatterji, “Segregation kinetics of topologically-modified ring polymers in cylin- drical confinement - Data, GitHub Repository,” https://github.com/harsh-doshi-729/Segregation-Kinetics- (2026)
work page 2026
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.