Invasion with size-dependent dispersion range
Pith reviewed 2026-07-01 01:58 UTC · model grok-4.3
The pith
Making dispersal range depend on colony size produces four distinct invasion growth regimes.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
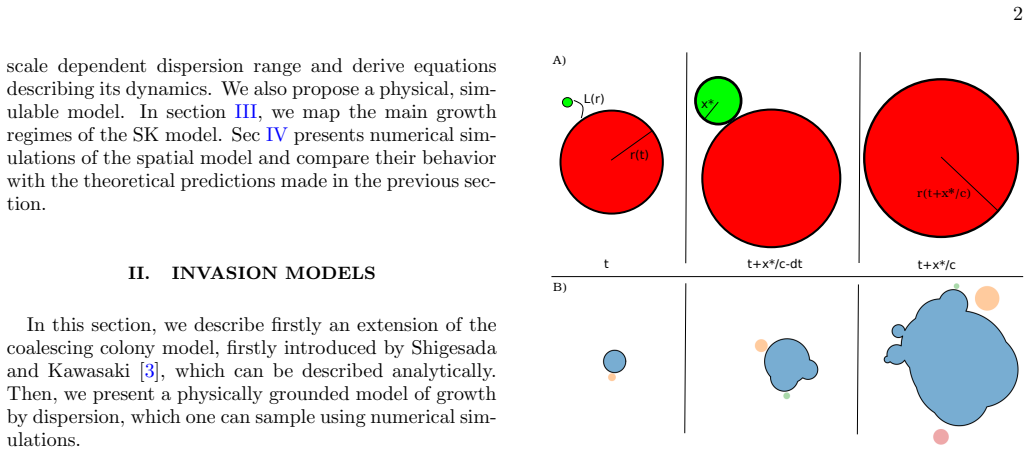
In the generalized coalescing colony model with size-dependent dispersion range r^μ, the main colony exhibits linear expansion, power-law growth, exponential regime, and finite-time blow-up depending on the value of μ. The coalescing colony approach correctly captures the scaling of the perimeter but fails to predict the scaling of the volume due to an effective breakdown of circular symmetry in the morphology of the main colony. The population fraction outside the main colony decays to zero like a power-law when μ<1, and a macroscopic amount remains in secondary colonies at μ=1.
What carries the argument
The size-dependent dispersion distance given by r^μ for a main colony of size r, which is used to derive the generalized dynamical equations and map the growth phase diagram.
Load-bearing premise
The main colony maintains circular symmetry allowing perimeter and volume scalings to be derived from the coalescing colony equations.
What would settle it
Measuring the actual volume growth exponent in a physical simulation for μ near 1 and checking if it matches the blow-up prediction or deviates due to shape irregularity.
Figures




read the original abstract
The coalescing colony model provides a minimal framework for biological invasions with long-range dispersion. In its standard formulation, the dispersion range is assumed independent of the size of the invading population. Here, we relax this assumption and consider size-dependent dispersal: a main colony of linear size $r$ emits secondary colonies at distance $r^\mu$, with $0 \leq \mu \leq 1$. We derive the generalized dynamical equations for this extended model and map out the growth phase diagram for the leading order contribution. Depending on $\mu$, the main colony exhibits distinct regimes: linear expansion, power-law growth, exponential regime and finite-time blow-up. We confront these theoretical predictions with a spatially explicit physical model. While the coalescing colony approach correctly captures the scaling of the perimeter, it fails to predict the scaling of the volume. We trace this discrepancy to an effective breakdown of circular symmetry in the morphology of the main colony. Finally, we quantify temporal evolution of the population fraction residing outside of the main colony. The coalescing colony model predicts its decay to~$0$ like a power-law when~$\mu<1$, and a macroscopic amount of the population remains in the secondary colonies at~$\mu=1$. Simulations of the physical model reveal a persistent satellite population not captured by the theory at~$\mu>\mu^*\approx 0.7$. Broadly, our findings highlight how coupling dispersal range to population size fundamentally alters invasion dynamics, with implications for biological invasions, metastatic growth, and urban expansion.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper extends the coalescing colony model to size-dependent dispersal, where a main colony of linear size r emits secondary colonies at distance r^μ (0 ≤ μ ≤ 1). It derives generalized dynamical equations, maps a growth phase diagram with μ-dependent regimes for the main colony (linear expansion, power-law growth, exponential regime, finite-time blow-up), compares predictions to spatially explicit simulations (perimeter scaling matches but volume does not, due to effective breakdown of circular symmetry), and quantifies the temporal decay of the population fraction outside the main colony (power-law decay for μ<1, macroscopic remainder at μ=1, with simulations showing persistent satellites for μ>μ*≈0.7).
Significance. If the derivations of the dynamical equations and phase diagram are robust, the work provides a clear extension of invasion models to size-dependent dispersal and identifies new dynamical regimes with potential implications for biological invasions and related processes. The explicit simulation comparison and attribution of the volume discrepancy to symmetry breakdown are useful for delineating model limitations.
major comments (3)
- [theoretical model / derivation of dynamical equations] Derivation of generalized dynamical equations (theoretical model section): the leading-order phase diagram for the main colony growth regimes appears to close the perimeter-to-volume relation and average long-range jumps under an implicit circular symmetry assumption; given that the physical model exhibits symmetry breakdown that already invalidates the volume scaling, it is necessary to show whether this assumption is required for the reported regimes (linear, power-law, exponential, blow-up) or whether they survive morphological fluctuations.
- [comparison with physical model] Comparison to simulations (results section): the claim that the coalescing colony approach 'correctly captures the scaling of the perimeter' while failing for volume is load-bearing for validating the phase diagram; however, no quantitative error bounds, symmetry measure (e.g., deviation from circularity), or sensitivity analysis is provided to assess how the observed morphological fluctuations propagate back into the predicted growth exponents or the location of regime boundaries.
- [population fraction outside main colony] Satellite population analysis: the prediction of power-law decay to zero for μ<1 versus macroscopic remainder at μ=1 is presented as a testable distinction, yet the reported simulation mismatch for μ>μ*≈0.7 is left without a proposed extension or parameter adjustment; this discrepancy directly tests the completeness of the leading-order closure and should be addressed to support the phase diagram's applicability.
minor comments (2)
- The abstract and main text should explicitly state the section or equation numbers where the generalized dynamical equations are derived and where the phase diagram is obtained, to allow readers to trace the symmetry assumptions.
- Figure captions for the simulation comparisons should include the number of realizations, error bars on the measured exponents, and the precise definition of 'volume' and 'perimeter' used in the physical model.
Simulated Author's Rebuttal
We thank the referee for the thoughtful and constructive report. We address the three major comments below, providing clarifications on the model's assumptions, agreeing where additional quantification is warranted, and outlining planned revisions.
read point-by-point responses
-
Referee: Derivation of generalized dynamical equations (theoretical model section): the leading-order phase diagram for the main colony growth regimes appears to close the perimeter-to-volume relation and average long-range jumps under an implicit circular symmetry assumption; given that the physical model exhibits symmetry breakdown that already invalidates the volume scaling, it is necessary to show whether this assumption is required for the reported regimes (linear, power-law, exponential, blow-up) or whether they survive morphological fluctuations.
Authors: The phase diagram is obtained from a leading-order closure that explicitly assumes a circular main colony to relate perimeter growth to the size-dependent dispersal distance r^μ. This assumption is required for the analytic derivation of the four regimes, as it allows averaging the long-range jumps and closing the equations for the effective radius. Simulations confirm that perimeter scaling (the quantity controlling the growth law) remains consistent with the circular prediction despite shape fluctuations, while volume deviates precisely because of the broken symmetry. We will revise the theoretical model section to state explicitly that the reported regimes apply under the circular approximation and note that strong morphological fluctuations could in principle shift regime boundaries, although the μ-dependent transitions in dispersal scaling are expected to persist at leading order. revision: partial
-
Referee: Comparison to simulations (results section): the claim that the coalescing colony approach 'correctly captures the scaling of the perimeter' while failing for volume is load-bearing for validating the phase diagram; however, no quantitative error bounds, symmetry measure (e.g., deviation from circularity), or sensitivity analysis is provided to assess how the observed morphological fluctuations propagate back into the predicted growth exponents or the location of regime boundaries.
Authors: We agree that quantitative support is needed to substantiate the perimeter agreement. In the revised manuscript we will add (i) a symmetry diagnostic (perimeter-to-equivalent-circle ratio averaged over runs), (ii) error bars on all fitted growth exponents obtained from multiple independent realizations, and (iii) a brief sensitivity check showing that the extracted exponents remain within the theoretically predicted regime for the observed range of shape fluctuations. These additions will directly address how morphological deviations affect the reported scalings. revision: yes
-
Referee: Satellite population analysis: the prediction of power-law decay to zero for μ<1 versus macroscopic remainder at μ=1 is presented as a testable distinction, yet the reported simulation mismatch for μ>μ*≈0.7 is left without a proposed extension or parameter adjustment; this discrepancy directly tests the completeness of the leading-order closure and should be addressed to support the phase diagram's applicability.
Authors: The observed persistence of satellites for μ ≳ 0.7 indicates that the single-main-colony closure becomes incomplete once secondary colonies reach comparable size and fail to coalesce rapidly. We will expand the discussion section to acknowledge this limitation of the leading-order model and sketch a possible multi-colony extension in which the largest colonies are tracked separately when their sizes exceed a threshold fraction of the main colony. A full quantitative treatment of the multi-colony regime lies beyond the present scope but is identified as a natural direction for future work. revision: partial
Circularity Check
No significant circularity; derivations are model extensions compared to independent simulations
full rationale
The paper derives generalized dynamical equations for size-dependent dispersal (r^μ) from the coalescing colony model extension and obtains phase diagram regimes (linear, power-law, exponential, blow-up). These are then tested against a separate spatially explicit physical model. The reported mismatch in volume scaling (due to symmetry breakdown) is explicitly noted rather than hidden, and perimeter agreement is presented as partial confirmation. No self-citations, fitted inputs renamed as predictions, or ansatz smuggling appear in the provided abstract or reader's summary. The central claims rest on explicit derivation plus external simulation comparison, making the work self-contained against benchmarks.
Axiom & Free-Parameter Ledger
free parameters (1)
- μ
axioms (1)
- domain assumption The coalescing colony model provides a minimal framework for biological invasions with long-range dispersion.
Reference graph
Works this paper leans on
-
[1]
The dispersal range isL(r) =L 0rµ, and for simplic- ity we setL 0 = 1. Dispersion exactly at rangeL(r) approximates more realistic situations where the dispersion range is range is stochastic, e.g. there is variance around an average dispersion range
-
[2]
The instantaneous absorption of colony is unreal- istic in most cases but helps for analytical predic- tions
-
[3]
The Shigesada-Kawasaki equations describe the growth of an average realisation. These equations are derived using a mean-field assumption: there is acontinousemission and absorption of colonies, which allows to write a density function for off- springs ρ(x, t) = ( 0 whenx > ct 1 c λ(r(t− x c )) otherwise. (7) The dynamics ofr(Eq. 4) can be written thanks ...
-
[4]
We focus our study on the caseµ≤1, that is dispersion occurs sublinearly or at most linearly. Beyond that limit, i.e. when the dispersion range scalessuper-linearly, the model loses consistency, as newly formed colonies would eventually disperse over distances larger than the primary one. B. Physical model Secondly, we consider thephysical model. The main...
-
[5]
R. J. Petit, E. Pineau, B. Demesure, R. Bacilieri, A. Ducousso, and A. Kremer, Proceedings of the National Academy of Sciences94, 9996 (1997). 8
work page 1997
-
[6]
J. S. Clark, M. Lewis, and L. Horvath, The American Naturalist157, 537 (2001), pMID: 18707261
work page 2001
-
[7]
N. Shigesada and K. Kawasaki, Invasion by stratified dif- fusion, inBiological invasions: theory and practice(Ox- ford University Press, USA, 1997) Chap. 5, p. 79–103
work page 1997
-
[8]
N. Shigesada, K. Kawasaki, and Y. Takeda, The Ameri- can Naturalist146, 229 (1995)
work page 1995
-
[9]
N. Shigesada and K. Kawasaki, Invasion and the range expansion of species: effects of long-distance dispersal, inDispersal Ecoogy. The42Symposium of the British Ecological Society(Blackwell Science, 2002) Chap. 17, p. 350–373
work page 2002
-
[10]
J. H. Arias, J. G´ omez-Garde˜ nes, S. Meloni, and E. Estrada, Journal of Theoretical Biology453, 1 (2018)
work page 2018
-
[11]
J. M. Bullock, L. Mallada Gonz´ alez, R. Tamme, L. G¨ otzenberger, S. M. White, M. P¨ artel, and D. A. P. Hooftman, Journal of Ecology105, 6 (2017)
work page 2017
-
[12]
J. S. Clark, The American Naturalist152, 204 (1998)
work page 1998
- [13]
-
[14]
V. Haustein and U. Schumacher, Journal of Clinical Bioinformatics2, 11 (2012)
work page 2012
- [15]
-
[16]
U. Marquis, O. Artime, R. Gallotti, and M. Barthelemy, Phys. Rev. Lett.135, 187403 (2025)
work page 2025
- [17]
-
[18]
R. A. Fisher, Annals of Eugenics7, 353 (1937)
work page 1937
-
[19]
A. N. Kolmogorov, I. G. Petrovskii, and N. Piskunov, Moscow University Bulletin of Mathematics1, 1 (1937)
work page 1937
-
[20]
U. Marquis, H. Berestycki, and M. Barthelemy, Euro- physics Letters (2026)
work page 2026
-
[21]
Beyond dynamic scaling: rare events break universality
U. Marquis, R. Gallotti, and M. Barthelemy, Beyond dynamic scaling: rare events break universality (2026), arXiv:2604.01820 [cond-mat.stat-mech]
work page internal anchor Pith review Pith/arXiv arXiv 2026
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.