A simple model of co-emergence of grid and place fields
Pith reviewed 2026-05-21 03:12 UTC · model grok-4.3
The pith
A recurrent network trained solely on predicting next sensory observations from masked inputs and motion develops both grid and place cells.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The central claim is that grid cells and place cells can co-emerge in a single recurrent network obeying Dale's Law when the network is trained to predict future sensory inputs from incomplete past sensory data plus self-motion information, providing a circuit-level account for their joint development during navigation.
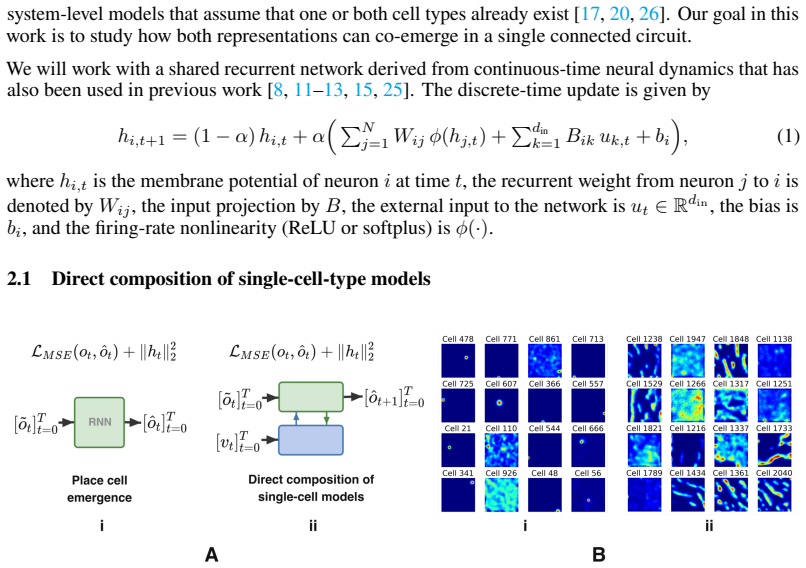
What carries the argument
A recurrent neural network with Dale's law constraint trained on a next-step sensory prediction task using masked observations and egocentric motion.
If this is right
- The relative numbers of grid-like and place-like units depend on the levels of sensory noise and input masking during training.
- The network reproduces grid fragmentation in hairpin mazes, merging of grids after wall removal, alignment of lattices across rooms, and ordered 3D fields in flying bats.
- Place cells develop before grid cells, matching the observed developmental timeline in animals.
- Two encoding pressures within the prediction objective explain the codes: one for reconstructing missing sensory parts and one for forecasting the next state during movement.
Where Pith is reading between the lines
- If the model holds, general predictive learning in sensory systems could naturally give rise to spatial maps without specialized spatial priors.
- Similar architectures might produce other functional cell types when trained on different prediction tasks.
- Experiments could test whether increasing sensory noise during animal navigation shifts the balance toward more place cells or grid cells.
Load-bearing premise
The masked sensory observations and egocentric motion used in training have the same statistical structure as what an animal experiences during natural exploration.
What would settle it
Training the network on sensory data or motion patterns that differ substantially from natural navigation and finding that neither grid nor place fields emerge would falsify the claim that the prediction objective alone suffices.
Figures


read the original abstract
Grid cells in the medial entorhinal cortex and place cells in the hippocampus together support spatial navigation. The two regions are reciprocally connected, and there is a chicken-and-egg problem for how both arise and reinforce each other during development. Current computational accounts either derive one type from the other or use network dynamics to model the emergence of one type in isolation. We introduce a unified recurrent network model that instantiates Dale's Law (every neuron is either excitatory or inhibitory), and is trained to predict the next sensory observation from masked previous sensory observations and egocentric motion. To our knowledge, this is the first single-objective model in which grid and place cells co-emerge without supervision of either type, or reliance on pre-existing spatial-cell representations. The two kinds of spatial codes coexist across 1,000 different training configurations, with their balance set by the amount of sensory noise and masking. Without retraining, the network qualitatively reproduces experimentally observed grid fragmentation in hairpin mazes, grid merging after wall removal, lattice alignment across connected rooms, locally ordered 3D fields observed in freely flying bats, as well as the developmental order in which place cells precede grid cells. We interpret these results in terms of two complementary encoding pressures within a single sensory-prediction objective: (1) correcting errors or reconstructing missing components of sensory observations, and (2) prediction of the next sensory state during navigation. Our results suggest a circuit-level account of the co-emergence of grid and place cells, and experimentally testable predictions for the two kinds of spatial codes.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces a recurrent neural network obeying Dale's law and trained solely to predict the next sensory observation from masked prior observations plus egocentric motion inputs. Grid-like and place-like codes emerge in the hidden units without explicit supervision on either; their relative prevalence is controlled by sensory noise and masking fraction. Across 1,000 training runs the model qualitatively reproduces grid fragmentation in hairpin mazes, grid merging after wall removal, lattice alignment across rooms, 3-D fields in flying bats, and the developmental precedence of place over grid cells.
Significance. If the central claim holds, the work supplies a single-objective, circuit-level account of how grid and place representations can co-emerge and mutually reinforce each other, directly addressing the chicken-and-egg problem. The reproduction of multiple distinct experimental signatures without retraining, the parameter sweep over 1,000 configurations, and the generation of falsifiable predictions constitute clear strengths.
major comments (1)
- [Methods (training-data generation)] Methods (training-data generation): the generative process for the sensory observation vector—its dimensionality, feature statistics, and dependence on true position versus raw egocentric signals—is not shown to be free of latent spatial structure. If observations are generated from a fixed 2-D layout containing position-dependent features, the network can solve the task by learning an implicit position embedding; this would render the result consistent with prior path-integration models rather than a pure demonstration that the prediction objective alone suffices.
minor comments (2)
- [Abstract and Results] Abstract and Results: quantitative metrics (e.g., gridness scores with error bars, place-field stability indices, or ablation controls removing recurrence or Dale's law) are absent; their addition would strengthen the claim that the observed codes arise robustly from the objective.
- [Figure captions and text] Figure captions and text: the precise definition of 'masking fraction' and how it interacts with sensory noise level across the 1,000 runs could be stated more explicitly to allow direct replication.
Simulated Author's Rebuttal
We thank the referee for their careful reading and for identifying this methodological point. We address the concern directly below and will revise the manuscript to strengthen the description of the training-data generation process.
read point-by-point responses
-
Referee: Methods (training-data generation): the generative process for the sensory observation vector—its dimensionality, feature statistics, and dependence on true position versus raw egocentric signals—is not shown to be free of latent spatial structure. If observations are generated from a fixed 2-D layout containing position-dependent features, the network can solve the task by learning an implicit position embedding; this would render the result consistent with prior path-integration models rather than a pure demonstration that the prediction objective alone suffices.
Authors: We agree that the current Methods section does not provide a sufficiently explicit account of the observation-generation process. In the model the sensory vector is produced by a fixed but locally observable feature map whose elements are sampled from a stationary distribution that depends only on the agent's instantaneous egocentric viewpoint; no global coordinate, absolute position tag, or pre-computed spatial embedding is ever supplied to the network. The only spatial information available to the recurrent units is therefore the egocentric velocity signal together with the history of masked observations that must be predicted. Nevertheless, we acknowledge that a reader could reasonably worry about hidden spatial structure, and we will add a dedicated subsection (with pseudocode and explicit parameter values) that (i) states the dimensionality and statistical independence of the feature channels, (ii) shows that each observation is generated solely from the current viewpoint without reference to a global map, and (iii) contrasts this construction with supervised path-integration models that receive explicit position targets. This revision will make the absence of latent spatial supervision unambiguous. revision: yes
Circularity Check
No significant circularity: spatial codes emerge from sensory-prediction objective without direct fitting or definitional reduction.
full rationale
The paper trains a recurrent network (instantiating Dale's law) solely on a next-observation prediction loss given masked sensory vectors plus egocentric velocity inputs. Grid and place fields are reported as post-training observations that arise as a byproduct of minimizing this loss across 1000 configurations, with balance controlled by noise/masking levels. No equation defines the fields in terms of each other or renames a fitted parameter as a prediction; the model does not optimize for spatial metrics. Experimental reproductions (hairpin fragmentation, 3D bat fields, developmental order) occur without retraining, confirming the outcome is not forced by construction. The input statistics assumption is a modeling premise but does not collapse the derivation chain to a tautology or self-citation load-bearing step. The central claim therefore remains independent of its inputs.
Axiom & Free-Parameter Ledger
free parameters (2)
- sensory noise level
- masking fraction
axioms (1)
- domain assumption Dale's Law: every neuron is strictly excitatory or strictly inhibitory
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
unified recurrent network model that instantiates Dale’s Law ... trained to predict the next sensory observation from masked previous sensory observations and egocentric motion
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
two complementary encoding pressures within a single sensory-prediction objective
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Torkel Hafting, Marianne Fyhn, Sturla Molden, May-Britt Moser, and Edvard I. Moser. Microstructure of a spatial map in the entorhinal cortex.Nature, 436(7052):801–806, August
-
[2]
ISSN 0028-0836, 1476-4687. doi: 10.1038/nature03721
-
[3]
Moser, Emilio Kropff, and May-Britt Moser
Edvard I. Moser, Emilio Kropff, and May-Britt Moser. Place Cells, Grid Cells, and the Brain’s SpatialRepresentationSystem.Annu.Rev.Neurosci.,31(1):69–89,July2008. ISSN0147-006X, 1545-4126. doi: 10.1146/annurev.neuro.31.061307.090723
-
[4]
A principle of economy predicts the functional architecture of grid cells.Elife, 4:e08362, 2015
Xue-Xin Wei, Jason Prentice, and Vijay Balasubramanian. A principle of economy predicts the functional architecture of grid cells.Elife, 4:e08362, 2015
work page 2015
-
[5]
J. O’Keefe and J. Dostrovsky. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat.Brain Research, 34(1):171–175, November 1971. ISSN 00068993. doi: 10.1016/0006-8993(71)90358-1
-
[6]
Placeunitsinthehippocampusofthefreelymovingrat.ExperimentalNeurology, 51(1):78–109, January 1976
JohnO’Keefe. Placeunitsinthehippocampusofthefreelymovingrat.ExperimentalNeurology, 51(1):78–109, January 1976. ISSN 00144886. doi: 10.1016/0014-4886(76)90055-8
-
[7]
May-Britt Moser, David C. Rowland, and Edvard I. Moser. Place Cells, Grid Cells, and Memory.Cold Spring Harb Perspect Biol, 7(2):a021808, February 2015. ISSN 1943-0264. doi: 10.1101/cshperspect.a021808
-
[8]
Yoram Burak and Ila R. Fiete. Accurate Path Integration in Continuous Attractor Network Models of Grid Cells.PLoS Comput Biol, 5(2):e1000291, February 2009. ISSN 1553-7358. doi: 10.1371/journal.pcbi.1000291
-
[9]
Ben Sorscher, Gabriel C. Mel, Samuel A. Ocko, Lisa M. Giocomo, and Surya Ganguli. A unified theory for the computational and mechanistic origins of grid cells.Neuron, 111(1): 121–137.e13, January 2023. ISSN 08966273. doi: 10.1016/j.neuron.2022.10.003
-
[10]
Kimberly L Stachenfeld, Matthew M Botvinick, and Samuel J Gershman. The hippocampus as a predictive map.Nat Neurosci, 20(11):1643–1653, November 2017. ISSN 1097-6256, 1546-1726. doi: 10.1038/nn.4650
-
[11]
Marcus K. Benna and Stefano Fusi. Place cells may simply be memory cells: Memory compressionleadstospatialtuningandhistorydependence.Proc.Natl.Acad.Sci.U.S.A.,118(51): e2018422118, December 2021. ISSN 0027-8424, 1091-6490. doi: 10.1073/pnas.2018422118
-
[12]
Di Tullio, Spencer Rooke, and Vijay Balasubramanian
Zhaoze Wang, Ronald W. Di Tullio, Spencer Rooke, and Vijay Balasubramanian. Time Makes Space: Emergence of Place Fields in Networks Encoding Temporally Continuous Sensory Experiences. InNeurIPS, 2024. doi: 10.1101/2024.08.11.607484
-
[13]
Andrea Banino, Caswell Barry, Benigno Uria, Charles Blundell, Timothy Lillicrap, Piotr Mirowski, Alexander Pritzel, Martin J. Chadwick, Thomas Degris, Joseph Modayil, Greg Wayne, Hubert Soyer, Fabio Viola, Brian Zhang, Ross Goroshin, Neil Rabinowitz, Razvan Pascanu, Charlie Beattie, Stig Petersen, Amir Sadik, Stephen Gaffney, Helen King, Koray Kavukcuoglu...
-
[14]
Christopher J. Cueva and Xue-Xin Wei. Emergence of grid-like representations by training recurrent neural networks to perform spatial localization, March 2018
work page 2018
-
[15]
Unfolding the Black Box of Recurrent Neural Networks for Path Integration
Tianhao Chu, Yuling Wu, Neil Burgess, Zilong Ji, and Si Wu. Unfolding the Black Box of Recurrent Neural Networks for Path Integration. InNeurIPS, 2025
work page 2025
-
[16]
On Conformal Isometry of Grid Cells: Learning Distance-Preserving Position Embedding, February 2025
Dehong Xu, Ruiqi Gao, Wen-Hao Zhang, Xue-Xin Wei, and Ying Nian Wu. On Conformal Isometry of Grid Cells: Learning Distance-Preserving Position Embedding, February 2025
work page 2025
-
[17]
Qiaorong S Yu, Zhaoze Wang, and Vijay Balasubramanian. When and where: A model hippocampal network unifies formation of time cells and place cells.bioRxiv, pages 2026–03, 2026
work page 2026
-
[18]
Trygve Solstad, Edvard I. Moser, and Gaute T. Einevoll. From grid cells to place cells: A mathematical model.Hippocampus, 16(12):1026–1031, December 2006. ISSN 1050-9631, 1098-1063. doi: 10.1002/hipo.20244. 11
-
[19]
EdmundT.Rolls,SimonM.Stringer,andThomasElliot. Entorhinalcortexgridcellscanmapto hippocampal place cells by competitive learning.Network: Computation in Neural Systems, 17 (4):447–465, January 2006. ISSN 0954-898X, 1361-6536. doi: 10.1080/09548980601064846
-
[20]
Grid cells require excitatory drive from the hippocampus.Nat Neurosci, 16(3):309–317, March 2013
Tora Bonnevie, Benjamin Dunn, Marianne Fyhn, Torkel Hafting, Dori Derdikman, John L Kubie, Yasser Roudi, Edvard I Moser, and May-Britt Moser. Grid cells require excitatory drive from the hippocampus.Nat Neurosci, 16(3):309–317, March 2013. ISSN 1097-6256, 1546-1726. doi: 10.1038/nn.3311
-
[21]
César Rennó-Costa and Adriano B.L. Tort. Place and Grid Cells in a Loop: Implications for Memory Function and Spatial Coding.J. Neurosci., 37(34):8062–8076, August 2017. ISSN 0270-6474, 1529-2401. doi: 10.1523/JNEUROSCI.3490-16.2017
-
[22]
Daniel Bush, Caswell Barry, and Neil Burgess. What do grid cells contribute to place cell firing?Trends in Neurosciences, 37(3):136–145, March 2014. ISSN 01662236. doi: 10.1016/j.tins.2013.12.003
-
[23]
Genela Morris and Dori Derdikman. The chicken and egg problem of grid cells and place cells.Trends in Cognitive Sciences, 27(2):125–138, February 2023. ISSN 13646613. doi: 10.1016/j.tics.2022.11.003
-
[24]
An oscillatory interference model of grid cell firing.Hippocampus, 17(9):801–812, September 2007
Neil Burgess, Caswell Barry, and John O’Keefe. An oscillatory interference model of grid cell firing.Hippocampus, 17(9):801–812, September 2007. ISSN 1050-9631, 1098-1063. doi: 10.1002/hipo.20327
-
[25]
Louis Kang and Vijay Balasubramanian. A geometric attractor mechanism for self-organization of entorhinal grid modules.eLife, 8, August 2019. doi: 10.7554/eLife.46687
-
[26]
Self-Supervised Learning of Representations for Space Generates Multi-Modular Grid Cells
RylanSchaeffer,TzuhsuanMa,SanmiKoyejo,MikailKhona,CristóbalEyzaguirre,andIlaRani Fiete. Self-Supervised Learning of Representations for Space Generates Multi-Modular Grid Cells. InNeurIPS, September 2023
work page 2023
-
[27]
SarthakChandra,SugandhaSharma,RishidevChaudhuri,andIlaFiete. Episodicandassociative memoryfromspatialscaffoldsinthehippocampus.Nature, 638(8051):739–751, February2025. ISSN 0028-0836, 1476-4687. doi: 10.1038/s41586-024-08392-y
-
[28]
Mikail Khona, Sarthak Chandra, and Ila Fiete. Global modules robustly emerge from local interactions and smooth gradients.Nature, 640(8057):155–164, April 2025. ISSN 0028-0836, 1476-4687. doi: 10.1038/s41586-024-08541-3
-
[29]
REMI: Reconstructing Episodic Memory During Internally Driven Path Planning
Zhaoze Wang, Genela Morris, Dori Derdikman, Pratik Chaudhari, and Vijay Balasubramanian. REMI: Reconstructing Episodic Memory During Internally Driven Path Planning. InNeurIPS, 2025
work page 2025
-
[30]
Misha V. Tsodyks, William E. Skaggs, Terrence J. Sejnowski, and Bruce L. McNaughton. ParadoxicalEffectsofExternalModulationofInhibitoryInterneurons.J.Neurosci.,17(11):4382– 4388, June 1997. ISSN 0270-6474, 1529-2401. doi: 10.1523/JNEUROSCI.17-11-04382.1997
-
[31]
CarlvanVreeswijkandHaimSompolinsky. Chaosinneuronalnetworkswithbalancedexcitatory and inhibitory activity.Science, 274(5293):1724–1726, 1996. Accessed: 29 Apr 2026
work page 1996
-
[32]
Dori Derdikman, Jonathan R Whitlock, Albert Tsao, Marianne Fyhn, Torkel Hafting, May- Britt Moser, and Edvard I Moser. Fragmentation of grid cell maps in a multicompartment environment.Nat Neurosci, 12(10):1325–1332, October 2009. ISSN 1097-6256, 1546-1726. doi: 10.1038/nn.2396
-
[33]
Francis Carpenter, Daniel Manson, Kate Jeffery, Neil Burgess, and Caswell Barry. Grid Cells Form a Global Representation of Connected Environments.Current Biology, 25(9):1176–1182, May 2015. ISSN 09609822. doi: 10.1016/j.cub.2015.02.037
-
[34]
Wills, Francesca Cacucci, Neil Burgess, and John O’Keefe
Tom J. Wills, Francesca Cacucci, Neil Burgess, and John O’Keefe. Development of the Hippocampal Cognitive Map in Preweanling Rats.Science, 328(5985):1573–1576, June 2010. ISSN 0036-8075, 1095-9203. doi: 10.1126/science.1188224
-
[35]
Rosamund F. Langston, James A. Ainge, Jonathan J. Couey, Cathrin B. Canto, Tale L. Bjerknes, Menno P. Witter, Edvard I. Moser, and May-Britt Moser. Development of the Spatial Representation System in the Rat.Science, 328(5985):1576–1580, June 2010. ISSN 0036-8075, 1095-9203. doi: 10.1126/science.1188210. 12
-
[36]
LaurenzMuessig,JonasHauser,ThomasJosephWills,andFrancescaCacucci. ADevelopmental Switch in Place Cell Accuracy Coincides with Grid Cell Maturation.Neuron, 86(5):1167–1173, June 2015. ISSN 08966273. doi: 10.1016/j.neuron.2015.05.011
-
[37]
Gily Ginosar, Johnatan Aljadeff, Yoram Burak, Haim Sompolinsky, Liora Las, and Nachum Ulanovsky. Locally ordered representation of 3D space in the entorhinal cortex.Nature, 596 (7872):404–409,August2021. ISSN0028-0836,1476-4687. doi: 10.1038/s41586-021-03783-x
-
[38]
Whatgridcellsconveyaboutratlocation.Journal of Neuroscience, 28(27):6858–6871, 2008
IlaRFiete,YoramBurak,andTedBrookings. Whatgridcellsconveyaboutratlocation.Journal of Neuroscience, 28(27):6858–6871, 2008
work page 2008
-
[39]
DanielBush,CaswellBarry,DanielManson,andNeilBurgess. UsingGridCellsforNavigation. Neuron, 87(3):507–520, August 2015. ISSN 08966273. doi: 10.1016/j.neuron.2015.07.006
-
[40]
The entorhinal grid map is discretized.Nature, 492(7427):72–78, 2012
Hanne Stensola, Tor Stensola, Trygve Solstad, Kristian Frøland, May-Britt Moser, and Edvard I Moser. The entorhinal grid map is discretized.Nature, 492(7427):72–78, 2012
work page 2012
-
[41]
Experience-dependent rescaling of entorhinal grids.Nature neuroscience, 10(6):682–684, 2007
Caswell Barry, Robin Hayman, Neil Burgess, and Kathryn J Jeffery. Experience-dependent rescaling of entorhinal grids.Nature neuroscience, 10(6):682–684, 2007
work page 2007
-
[42]
Tor Stensola, Hanne Stensola, May-Britt Moser, and Edvard I. Moser. Shearing-induced asymmetry in entorhinal grid cells.Nature, 518(7538):207–212, February 2015. ISSN 0028-0836, 1476-4687. doi: 10.1038/nature14151
-
[43]
Grid cell symmetry is shaped by environmental geometry.Nature, 518(7538):232–235, February 2015
Julija Krupic, Marius Bauza, Stephen Burton, Caswell Barry, and John O’Keefe. Grid cell symmetry is shaped by environmental geometry.Nature, 518(7538):232–235, February 2015. ISSN 0028-0836, 1476-4687. doi: 10.1038/nature14153
-
[44]
Environmental boundaries as an error correction mechanism for grid cells.Neuron, 86(3):827–839, 2015
Kiah Hardcastle, Surya Ganguli, and Lisa M Giocomo. Environmental boundaries as an error correction mechanism for grid cells.Neuron, 86(3):827–839, 2015
work page 2015
-
[45]
Alexandra T Keinath, Russell A Epstein, and Vijay Balasubramanian. Environmental deforma- tions dynamically shift the grid cell spatial metric.eLife, 7:e38169, oct 2018. ISSN 2050-084X. doi: 10.7554/eLife.38169. URLhttps://doi.org/10.7554/eLife.38169
-
[46]
Francesca Sargolini, Marianne Fyhn, Torkel Hafting, Bruce L McNaughton, Menno P Witter, May-Britt Moser, and Edvard I Moser. Conjunctive representation of position, direction, and velocity in entorhinal cortex.Science, 312(5774):758–762, 2006. 13 S Supplementary material S.1 Continuous-time RNN dynamics The discrete-time update in Eq. 1 is derived from th...
work page 2006
-
[47]
The agent is spawned and confined to the left compartment
Room 0 (left sub-room). The agent is spawned and confined to the left compartment
-
[48]
The agent is respawned in the right compartment
Room 1 (right sub-room). The agent is respawned in the right compartment
-
[49]
The internal wall is removed, and the agent is allowed to move freely through the full arena
Merged arena. The internal wall is removed, and the agent is allowed to move freely through the full arena. At initialization, we first sample a random Gaussian noise field. We then generate the sensory response maps for the two rooms using the boundary-aware diffusion smoothing method described in Section S.3. Because this diffusion process respects envi...
work page 2048
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.