Experimental Collapse in Virophysics: Protocol-Resolved Observation, Inference, and Plaque-Assay Blindness
Pith reviewed 2026-06-29 09:16 UTC · model grok-4.3
The pith
Virological measurements are protocol-conditioned projections of a richer latent virion-environment ensemble.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
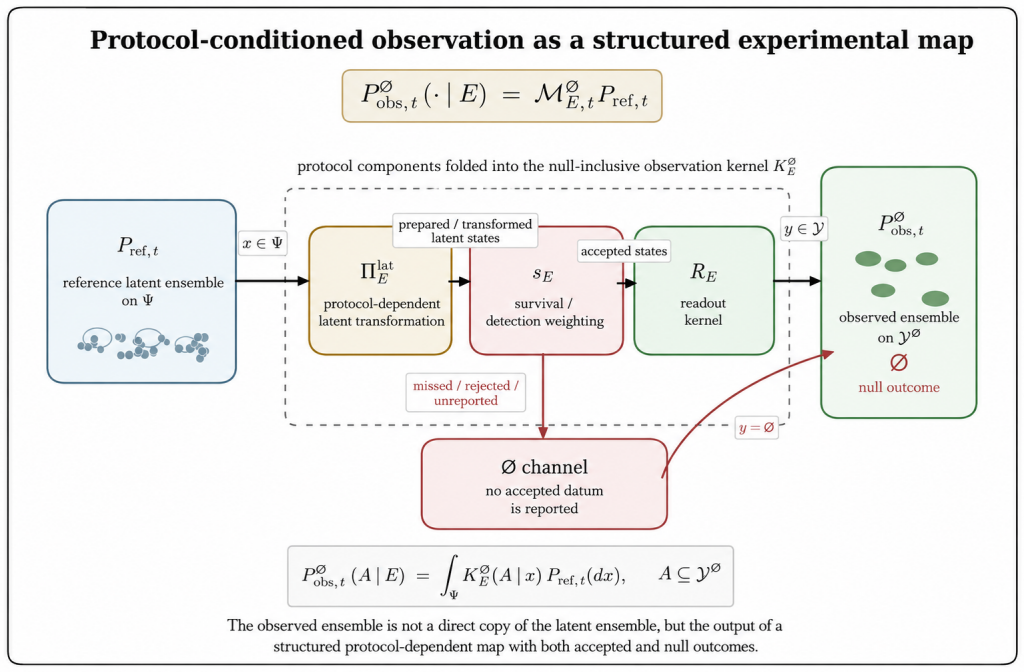
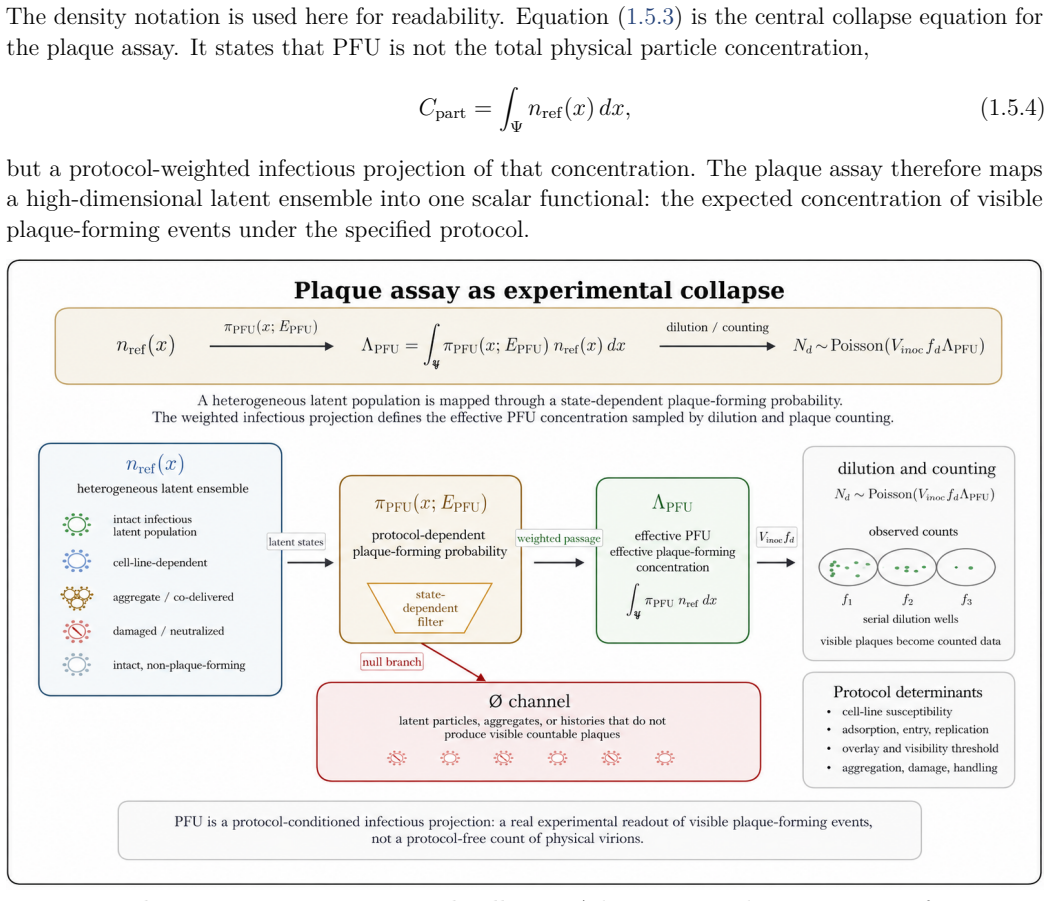
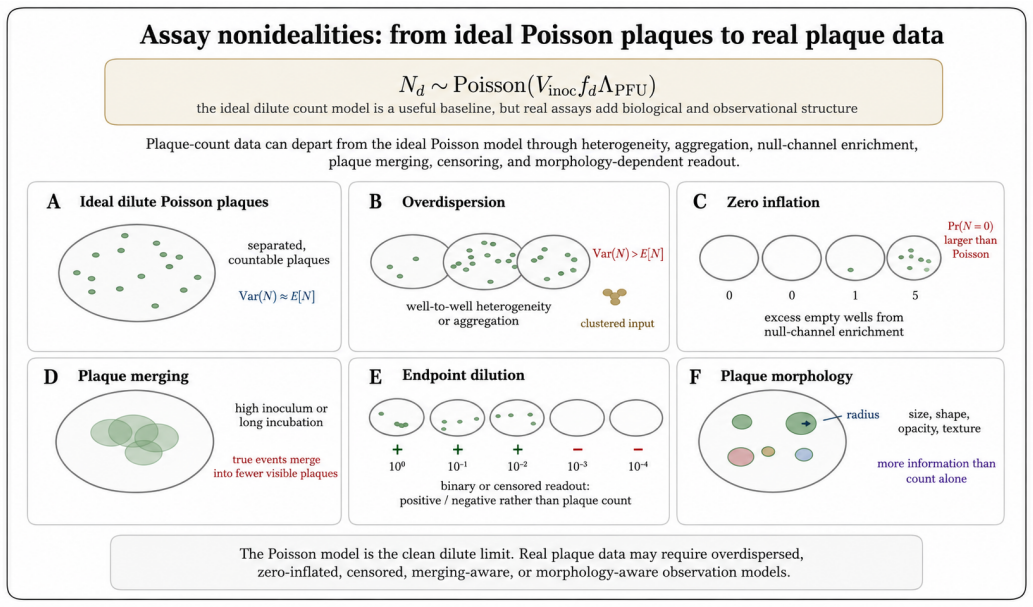
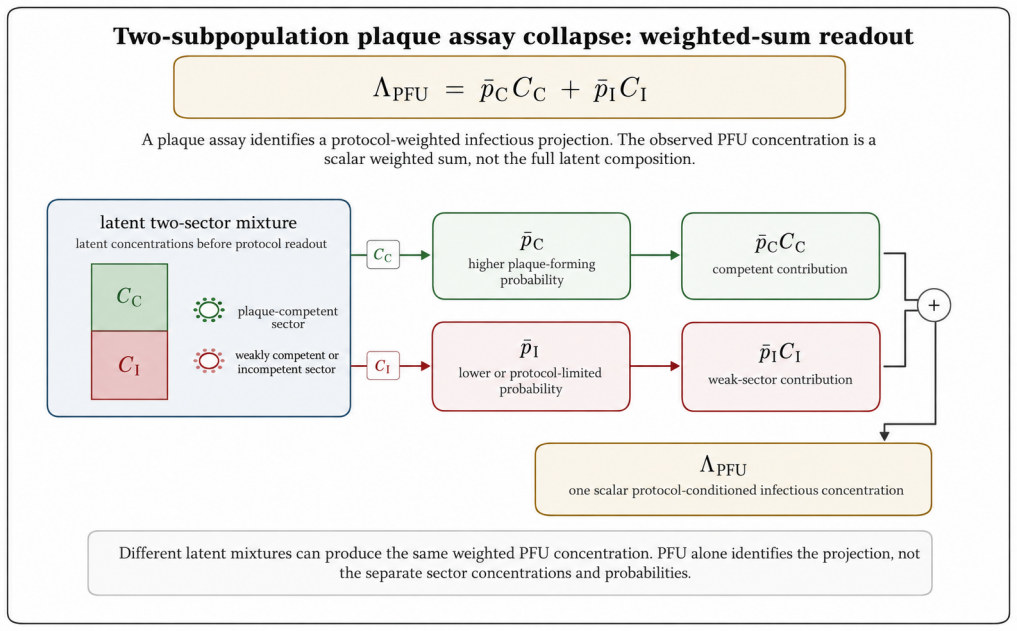
The paper claims that an experiment observes a protocol-conditioned projection of a richer latent virion-environment ensemble, formalized by the null-inclusive observation operator P_obs,t^∅(·|E) = M_E,t^∅ P_ref,t. This operator separates latent-state transformation, detection weighting, readout, and non-observation, making protocol effects explicit. The plaque assay therefore estimates an effective protocol-conditioned infectious concentration Λ_PFU = ∫ π_PFU(x; E_PFU) n_ref(x) dx rather than total particle concentration, recovering the Poisson plaque-count model in the dilute limit.
What carries the argument
The null-inclusive observation operator P_obs,t^∅(·|E) = M_E,t^∅ P_ref,t, which maps a reference latent ensemble to the observed ensemble generated by protocol E, including null outcomes.
If this is right
- The plaque assay estimates an effective protocol-conditioned infectious concentration rather than total particle concentration.
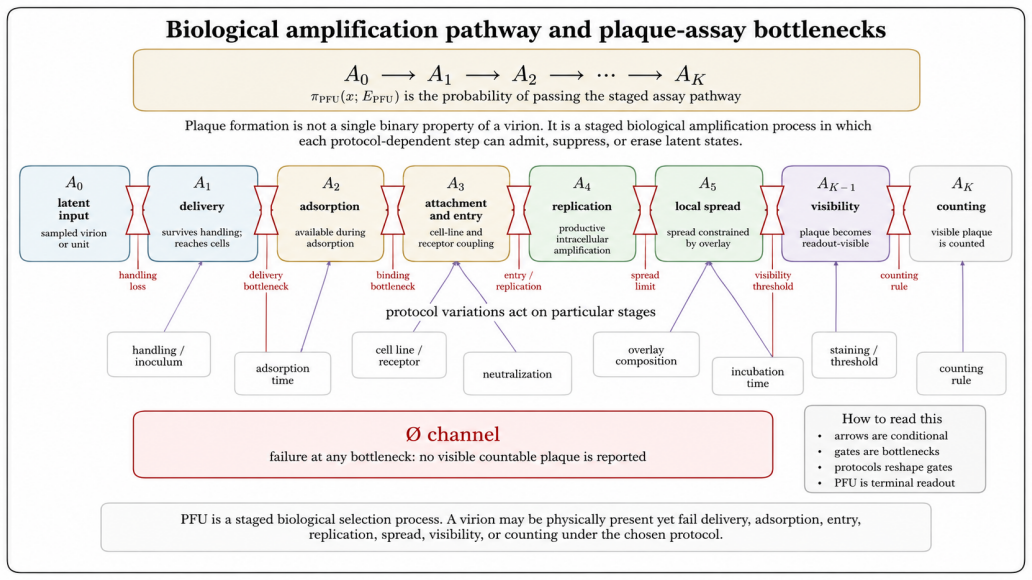
- Deviations such as overdispersion, zero inflation, plaque merging, and morphology variations are recast as protocol-conditioned information rather than noise.
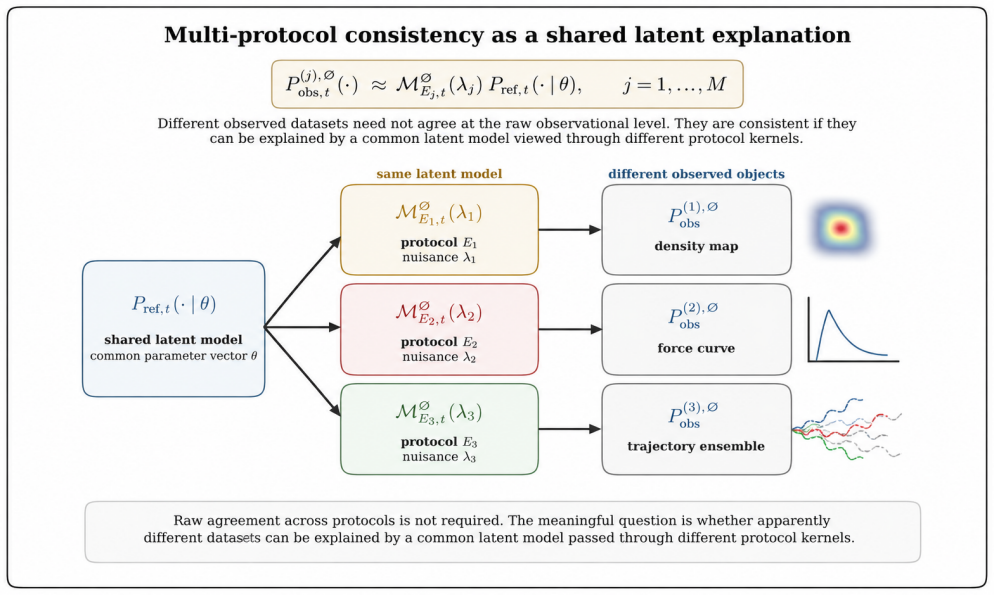
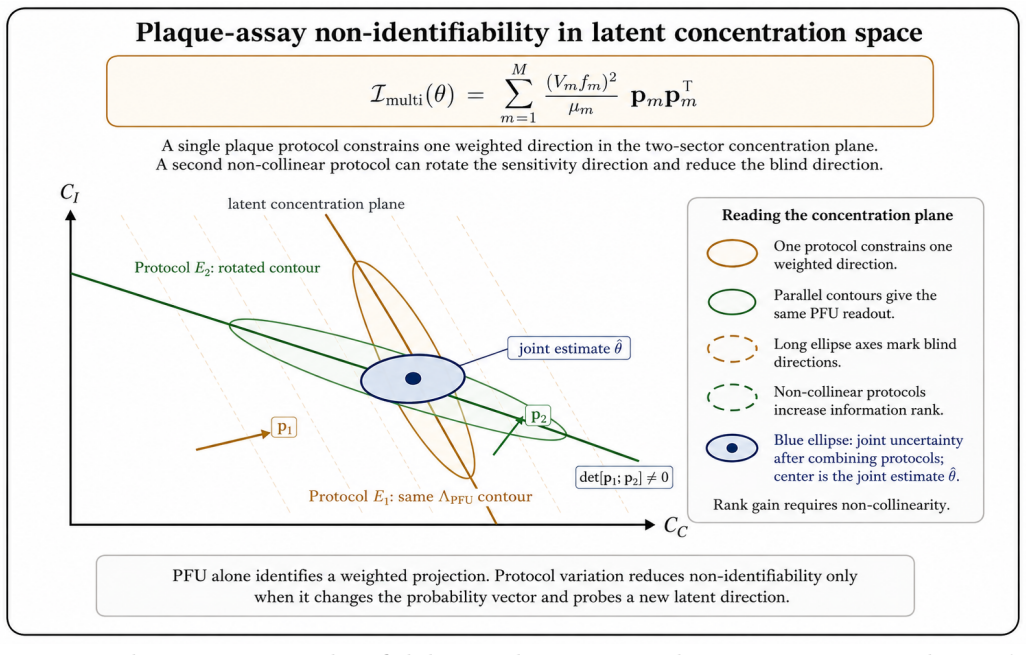
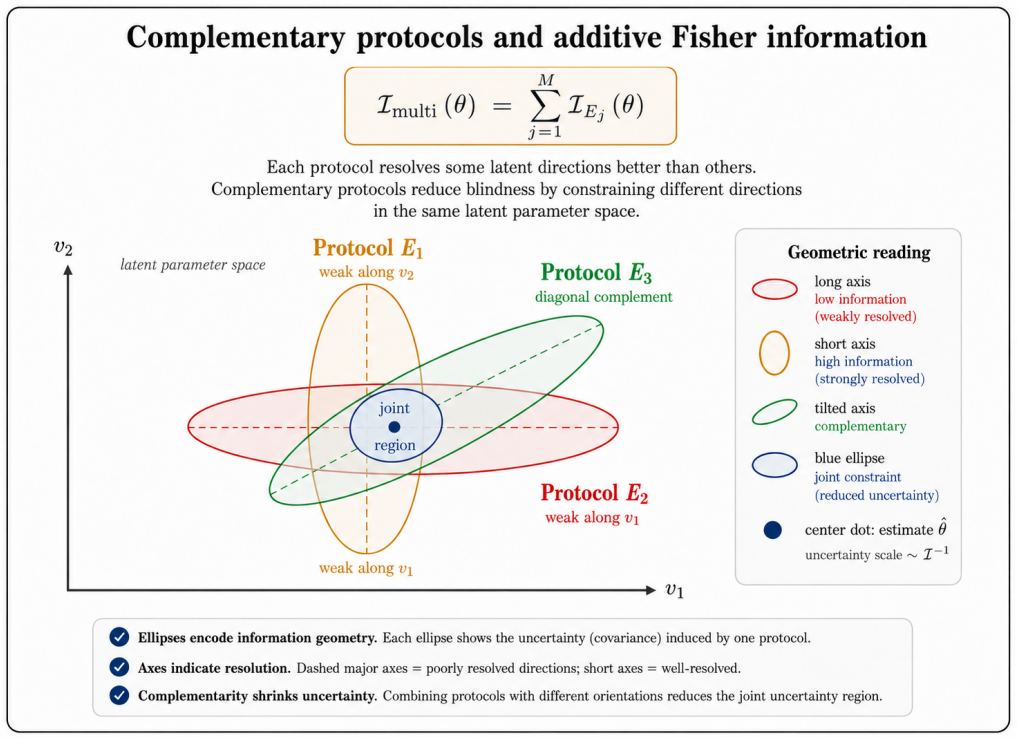
- Multi-protocol consistency checks allow inverse inference of hidden features in the reference latent ensemble.
- Complementary assays can be designed to cover different parts of the latent space.
Where Pith is reading between the lines
- The same operator decomposition could be applied to measurements in other complex systems where protocol choices shape what is observed.
- Protocols with deliberately complementary blind spots might be combined to reconstruct more of the latent ensemble than any single assay reveals.
- Direct tests of operator separability could be performed by varying one protocol component while holding others fixed and checking for independent effects on the output distribution.
Load-bearing premise
A richer latent virion-environment ensemble exists independently of any protocol and the effects of preparation, immobilization, loading, steering, filtering, amplification, censoring, and detection can be cleanly separated into distinct operator components.
What would settle it
An experiment in which two protocols cannot be decomposed into independent operator components because their effects on the latent ensemble are inseparably coupled.
Figures
read the original abstract
Virological measurements are often treated as reports of virion structure, mechanics, dielectric response, infectivity, or titer. In practice, an experiment observes a protocol-conditioned projection of a richer latent virion--environment ensemble. This paper defines this process as experimental collapse within protocol-resolved virophysics. Its central object is the null-inclusive observation operator $P_{\mathrm{obs},t}^{\varnothing}(\,\cdot\mid E\,) = \mathcal{M}_{E,t}^{\varnothing}P_{\mathrm{ref},t}$, which maps a reference latent ensemble to the observed ensemble generated by protocol $E$, including null outcomes. The formulation separates latent-state transformation, detection weighting, readout, and non-observation, making protocol effects explicit components rather than bias terms. The framework introduces protocol-conditioned latent ensembles, collapse functionals, protocol blindness, observation equivalence, Fisher-information observability, inverse inference, and multi-protocol consistency. It identifies collapse mechanisms including preparation, surface immobilization, mechanical loading, field steering, medium filtering, amplification, censoring, and detection thresholds. As a worked example, the plaque assay estimates an effective protocol-conditioned infectious concentration $\Lambda_{\mathrm{PFU}}=\int_{\Psi}\pi_{\mathrm{PFU}}(x;E_{\mathrm{PFU}})n_{\mathrm{ref}}(x),dx$, rather than total particle concentration. This recovers the Poisson plaque-count model and PFU titer formula in the dilute regime; extensions to overdispersion, zero inflation, plaque merging, endpoint dilution, neutralization, and morphology-augmented readouts recast deviations as protocol-conditioned information. Thus, virological data are outputs of explicit protocol kernels, clarifying what measurements report, miss, and how complementary assays can infer hidden latent virion structures.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript defines 'experimental collapse' in virophysics via the null-inclusive observation operator P_obs,t^∅(·|E) = M_E,t^∅ P_ref,t, which maps a reference latent virion-environment ensemble to a protocol-conditioned observed ensemble. It factors protocol effects (preparation, immobilization, mechanical loading, filtering, amplification, censoring, detection thresholds) into components of M, introduces related concepts such as protocol blindness and Fisher-information observability, and shows that the plaque-assay case recovers the standard Poisson plaque-count model and PFU titer formula in the dilute regime, with extensions to overdispersion and other deviations treated as protocol-conditioned information.
Significance. If the clean factorization of coupled physical mechanisms into independent operator components can be justified and tested, the framework would offer a systematic language for interpreting what virological assays report versus miss and for designing multi-protocol inference. The plaque-assay reduction is a consistency check rather than an independent prediction. No new empirical tests, parameter-free derivations, or falsifiable predictions beyond re-deriving known results are supplied.
major comments (2)
- [Abstract] Abstract and worked-example section: the claim that the operator cleanly separates latent transformation, detection weighting, readout, and non-observation rests on the axiom that the listed mechanisms (preparation, immobilization, mechanical loading, field steering, medium filtering, amplification, censoring, detection thresholds) act as independent factors. The manuscript supplies no argument or test showing that this factorization remains unique or information-preserving when mechanisms are physically coupled (e.g., surface immobilization simultaneously altering local concentration and detection thresholds).
- [Abstract] Abstract, Eq. for Λ_PFU: the plaque-assay reduction recovers the standard Poisson PFU formula by direct substitution once the protocol kernel π_PFU(x; E_PFU) is posited; this is a restatement of the definitional setup rather than an independent derivation or empirical validation of the broader inference claims.
minor comments (1)
- Notation for the observation operator and collapse functionals is introduced without an explicit comparison table to existing measurement models in virology, which would aid readability.
Simulated Author's Rebuttal
We thank the referee for the careful reading and for identifying the key modeling assumptions in the framework. We respond to each major comment below.
read point-by-point responses
-
Referee: [Abstract] Abstract and worked-example section: the claim that the operator cleanly separates latent transformation, detection weighting, readout, and non-observation rests on the axiom that the listed mechanisms (preparation, immobilization, mechanical loading, field steering, medium filtering, amplification, censoring, detection thresholds) act as independent factors. The manuscript supplies no argument or test showing that this factorization remains unique or information-preserving when mechanisms are physically coupled (e.g., surface immobilization simultaneously altering local concentration and detection thresholds).
Authors: We agree that the factorization is introduced as a modeling axiom to render protocol effects explicit rather than as a derived or empirically validated property. The manuscript does not supply a uniqueness proof or tests for coupled mechanisms, as the operator is offered as a conceptual language for structuring inference. When physical coupling occurs the decomposition may lose uniqueness, yet the composite mapping P_obs remains well-defined. We will add an explicit caveat on this assumption and its scope in a revised discussion section. revision: partial
-
Referee: [Abstract] Abstract, Eq. for Λ_PFU: the plaque-assay reduction recovers the standard Poisson PFU formula by direct substitution once the protocol kernel π_PFU(x; E_PFU) is posited; this is a restatement of the definitional setup rather than an independent derivation or empirical validation of the broader inference claims.
Authors: The plaque-assay case is presented precisely as a consistency check that recovers the known Poisson model and PFU formula upon substitution of the protocol kernel. This is not offered as an independent empirical validation or new derivation but as an illustration that the general formalism specializes correctly to an established result. The broader contribution concerns the utility of protocol-conditioned ensembles and blindness concepts for interpreting deviations in other regimes. revision: no
Circularity Check
Central observation operator introduced by definition; plaque-assay example recovers known PFU formula by construction
specific steps
-
self definitional
[Abstract]
"Its central object is the null-inclusive observation operator $P_{\mathrm{obs},t}^{\varnothing}(\,\cdot\mid E\,) = \mathcal{M}_{E,t}^{\varnothing}P_{\mathrm{ref},t}$, which maps a reference latent ensemble to the observed ensemble generated by protocol $E$, including null outcomes. The formulation separates latent-state transformation, detection weighting, readout, and non-observation, making protocol effects explicit components rather than bias terms."
The operator is defined to equal the product of M and P_ref; the claimed separation into distinct components is therefore true by the definition itself rather than derived from any independent physical argument or measurement.
-
fitted input called prediction
[Abstract]
"As a worked example, the plaque assay estimates an effective protocol-conditioned infectious concentration $\Lambda_{\mathrm{PFU}}=\int_{\Psi}\pi_{\mathrm{PFU}}(x;E_{\mathrm{PFU}})n_{\mathrm{ref}}(x),dx$, rather than total particle concentration. This recovers the Poisson plaque-count model and PFU titer formula in the dilute regime"
Once the protocol kernel $\pi_{\mathrm{PFU}}$ is inserted into the already-defined observation operator, the integral is constructed to reproduce the standard Poisson plaque-count and PFU formulas; the 'recovery' is therefore a restatement of the definitional setup rather than an independent prediction.
full rationale
The paper defines its central object as the composition P_obs = M P_ref and then presents the plaque-assay reduction as recovering the standard Poisson/PFU model once the protocol kernel is inserted. Both steps are tautological under the posited factorization; no independent derivation or external constraint is shown. The framework therefore restates its definitional setup rather than deriving new predictions from first principles.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption A richer latent virion-environment ensemble exists independently of measurement protocols.
- ad hoc to paper Protocol effects (preparation, immobilization, filtering, detection thresholds) can be cleanly factored into separate components of the operator.
invented entities (2)
-
experimental collapse
no independent evidence
-
protocol blindness
no independent evidence
Reference graph
Works this paper leans on
-
[1]
Single-particle cryo-EM at crystallographic resolution,
Y. Cheng, “Single-particle cryo-EM at crystallographic resolution,”Cell, vol. 161, no. 3, pp. 450–457,
-
[2]
doi: 10.1016/j.cell.2015.03.049
-
[3]
R. F. Thompson, M. Walker, C. A. Siebert, S. P. Muench, and N. A. Ranson, “An introduction to sample preparation and imaging by cryo-electron microscopy for structural biology,”Methods, vol. 100, pp. 3–15, 2016. doi: 10.1016/j.ymeth.2016.02.017
-
[4]
Reducing effects of particle adsorption to the air–water interface in cryo-EM,
A. J. Noble, H. Wei, V. P. Dandey, Z. Zhang, Y. Z. Tan, C. S. Potter, and B. Carragher, “Reducing effects of particle adsorption to the air–water interface in cryo-EM,”Nature Methods, vol. 15, pp. 793–795, 2018. doi: 10.1038/s41592-018-0139-3
-
[5]
J. Chen, A. J. Noble, J. Y. Kang, and S. A. Darst, “Eliminating effects of particle adsorp- tion to the air/water interface in single-particle cryo-electron microscopy: Bacterial RNA polymerase and CHAPSO,”Journal of Structural Biology: X, vol. 1, p. 100005, 2019. doi: 10.1016/j.yjsbx.2019.100005
-
[6]
Better cryo-EM specimen preparation: How to deal with the air–water interface?
N. Liu and H.-W. Wang, “Better cryo-EM specimen preparation: How to deal with the air–water interface?”Journal of Molecular Biology, vol. 435, no. 9, p. 167926, 2023. doi: 10.1016/j.jmb.2022.167926
-
[7]
Mechanical properties of viruses analyzed by atomic force microscopy: A virological perspective,
M. G. Mateu, “Mechanical properties of viruses analyzed by atomic force microscopy: A virological perspective,”Virus Research, vol. 168, no. 1–2, pp. 1–22, 2012. doi: 10.1016/j.virusres.2012.06.008
-
[8]
M. Marchetti, G. J. L. Wuite, and W. H. Roos, “Atomic force microscopy observation and characterization of single virions and virus-like particles by nano-indentation,”Current Opinion in Virology, vol. 18, pp. 82–88, 2016. doi: 10.1016/j.coviro.2016.05.002
-
[9]
Topography, spike dynamics, and nanome- chanics of individual native SARS-CoV-2 virions,
B. Kiss, Z. Kis, B. Pályi, and M. S. Z. Kellermayer, “Topography, spike dynamics, and nanome- chanics of individual native SARS-CoV-2 virions,”Nano Letters, vol. 21, no. 6, pp. 2675–2680,
-
[10]
doi: 10.1021/acs.nanolett.0c04465
-
[11]
Atomic force microscopy analysis of native infectious and inactivated SARS-CoV-2 virions,
S. Lyonnais, M. Hénaut, A. Neyret, P. Merida, C. Cazevieille, N. Gros, C. Chable-Bessia, and D. Muriaux, “Atomic force microscopy analysis of native infectious and inactivated SARS-CoV-2 virions,”Scientific Reports, vol. 11, article 11885, 2021. doi: 10.1038/s41598-021-91371-4. 170
-
[12]
Manipulation of herpes simplex virus type 1 by dielectrophoresis,
M. P. Hughes, H. Morgan, F. J. Rixon, J. P. H. Burt, and R. Pethig, “Manipulation of herpes simplex virus type 1 by dielectrophoresis,”Biochimica et Biophysica Acta: General Subjects, vol. 1425, no. 1, pp. 119–126, 1998. doi: 10.1016/S0304-4165(98)00058-0
-
[13]
Measuring the dielectric properties of herpes simplex virus type 1 virions with dielectrophoresis,
M. P. Hughes, H. Morgan, and F. J. Rixon, “Measuring the dielectric properties of herpes simplex virus type 1 virions with dielectrophoresis,”Biochimica et Biophysica Acta: General Subjects, vol. 1571, no. 1, pp. 1–8, 2002. doi: 10.1016/S0304-4165(02)00161-7
-
[14]
Review article—Dielectrophoresis: Status of the theory, technology, and applications,
R. Pethig, “Review article—Dielectrophoresis: Status of the theory, technology, and applications,” Biomicrofluidics, vol. 4, no. 2, p. 022811, 2010. doi: 10.1063/1.3456626
-
[15]
Dielectrophoresis: From molecular to micrometer-scale analytes,
D. Kim, M. Sonker, and A. Ros, “Dielectrophoresis: From molecular to micrometer-scale analytes,” Analytical Chemistry, vol. 91, no. 1, pp. 277–295, 2019. doi: 10.1021/acs.analchem.8b05454
-
[16]
Movements of HIV-virions in human cervical mucus,
H. Boukari, B. Brichacek, P. Stratton, S. F. Mahoney, J. D. Lifson, L. Margolis, and R. Nossal, “Movements of HIV-virions in human cervical mucus,”Biomacromolecules, vol. 10, no. 9, pp. 2482– 2488, 2009. doi: 10.1021/bm900344q
-
[17]
Influenza A virus diffusion through mucus gel networks,
L. Kaler, E. Iverson, S. Bader, D. Song, M. A. Scull, and G. A. Duncan, “Influenza A virus diffusion through mucus gel networks,”Communications Biology, vol. 5, article 249, 2022. doi: 10.1038/s42003-022-03204-3
-
[18]
Production of plaques in monolayer tissue cultures by single particles of an animal virus,
R. Dulbecco, “Production of plaques in monolayer tissue cultures by single particles of an animal virus,”Proceedings of the National Academy of Sciences of the United States of America, vol. 38, no. 8, pp. 747–752, 1952. doi: 10.1073/pnas.38.8.747
-
[19]
The plaque assay of animal viruses,
P. D. Cooper, “The plaque assay of animal viruses,”Advances in Virus Research, vol. 8, pp. 319–378,
-
[20]
doi: 10.1016/S0065-3527(08)60689-2
-
[21]
A. Baer and K. Kehn-Hall, “Viral concentration determination through plaque assays: Using traditional and novel overlay systems,”Journal of Visualized Experiments, no. 93, e52065, 2014. doi: 10.3791/52065
-
[22]
S. M. Kay,Fundamentals of Statistical Signal Processing, Volume I: Estimation Theory. Upper Saddle River, NJ: Prentice Hall PTR, 1993
1993
-
[23]
A. W. van der Vaart,Asymptotic Statistics. Cambridge: Cambridge University Press, 1998. doi: 10.1017/CBO9780511802256
-
[24]
T. M. Cover and J. A. Thomas,Elements of Information Theory, 2nd ed. Hoboken, NJ: Wiley- Interscience, 2006. doi: 10.1002/047174882X
-
[25]
Villani,Optimal Transport: Old and New
C. Villani,Optimal Transport: Old and New. Berlin: Springer, 2009. doi: 10.1007/978-3-540-71050- 9. 171
-
[26]
Mucus structure, viscoelastic properties, and composition in chronic respiratory diseases,
M. Abrami, A. Biasin, F. Tescione, D. Tierno, B. Dapas, A. Carbone, G. Grassi, M. Conese, S. Di Gioia, D. Larobina, and M. Grassi, “Mucus structure, viscoelastic properties, and composition in chronic respiratory diseases,”International Journal of Molecular Sciences, vol. 25, no. 3, article 1933, 2024. doi: 10.3390/ijms25031933
-
[27]
Influenza A virus surface proteins are organized to help penetrate host mucus,
M. D. Vahey and D. A. Fletcher, “Influenza A virus surface proteins are organized to help penetrate host mucus,”eLife, vol. 8, e43764, 2019. doi: 10.7554/eLife.43764
-
[28]
On information and sufficiency,
S. Kullback and R. A. Leibler, “On information and sufficiency,”The Annals of Mathematical Statistics, vol. 22, no. 1, pp. 79–86, 1951. doi: 10.1214/aoms/1177729694
-
[29]
Divergence measures based on the Shannon entropy,
J. Lin, “Divergence measures based on the Shannon entropy,”IEEE Transactions on Information Theory, vol. 37, no. 1, pp. 145–151, 1991. doi: 10.1109/18.61115
-
[30]
A kernel two-sample test,
A. Gretton, K. M. Borgwardt, M. J. Rasch, B. Schölkopf, and A. Smola, “A kernel two-sample test,”Journal of Machine Learning Research, vol. 13, no. 25, pp. 723–773, 2012
2012
-
[31]
Tarantola,Inverse Problem Theory and Methods for Model Parameter Estimation
A. Tarantola,Inverse Problem Theory and Methods for Model Parameter Estimation. Philadelphia, PA: Society for Industrial and Applied Mathematics, 2005. doi: 10.1137/1.9780898717921
-
[32]
Kaipio and E
J. Kaipio and E. Somersalo,Statistical and Computational Inverse Problems. New York: Springer,
-
[33]
doi: 10.1007/b138659
-
[34]
On structural identifiability,
R. Bellman and K. J. Åström, “On structural identifiability,”Mathematical Biosciences, vol. 7, no. 3–4, pp. 329–339, 1970. doi: 10.1016/0025-5564(70)90132-X
-
[35]
E. Walter and L. Pronzato,Identification of Parametric Models from Experimental Data. London: Springer, 1997. doi: 10.1007/978-3-642-58923-3
-
[36]
A. Raue, C. Kreutz, T. Maiwald, J. Bachmann, M. Schilling, U. Klingmüller, and J. Tim- mer, “Structural and practical identifiability analysis of partially observed dynamical models by exploiting the profile likelihood,”Bioinformatics, vol. 25, no. 15, pp. 1923–1929, 2009. doi: 10.1093/bioinformatics/btp358
-
[37]
Protein denaturation at the air–water interface and how to prevent it,
E. D’Imprima, D. Floris, M. Joppe, R. Sánchez, M. Grininger, and W. Kühlbrandt, “Protein denaturation at the air–water interface and how to prevent it,”eLife, vol. 8, e42747, 2019. doi: 10.7554/eLife.42747
-
[38]
T. S. Levitz, E. J. Brignole, I. Fong, M. C. Darrow, and C. L. Drennan, “Effects of chameleon dispense-to-plunge speed on particle concentration, complex formation, and final resolution: A case study using theNeisseria gonorrhoeaeribonucleotide reductase inactive complex,”Journal of Structural Biology, vol. 214, no. 1, p. 107825, 2022. doi: 10.1016/j.jsb....
-
[39]
Influenza-binding antibodies immobilise influenza viruses in fresh human airway mucus,
Y.-Y. Wang, D. Harit, D. B. Subramani, H. Arora, P. A. Kumar, and S. K. Lai, “Influenza-binding antibodies immobilise influenza viruses in fresh human airway mucus,”European Respiratory Journal, vol. 49, no. 1, p. 1601709, 2017. doi: 10.1183/13993003.01709-2016. 172
-
[40]
Plaque formation and isolation of pure lines with poliomyelitis viruses,
R. Dulbecco and M. Vogt, “Plaque formation and isolation of pure lines with poliomyelitis viruses,” Journal of Experimental Medicine, vol. 99, no. 2, pp. 167–182, 1954. doi: 10.1084/jem.99.2.167
-
[41]
Molecular determinants of the ratio of inert to infectious virus particles,
P. J. Klasse, “Molecular determinants of the ratio of inert to infectious virus particles,” Progress in Molecular Biology and Translational Science, vol. 129, pp. 285–326, 2015. doi: 10.1016/bs.pmbts.2014.10.012
-
[42]
Biological activities of ‘noninfectious’ influenza A virus particles,
C. B. Brooke, “Biological activities of ‘noninfectious’ influenza A virus particles,”Future Virology, vol. 9, no. 1, pp. 41–51, 2014. doi: 10.2217/fvl.13.118
-
[43]
Collective properties of viral infectivity,
R. Sanjuán, “Collective properties of viral infectivity,”Current Opinion in Virology, vol. 33, pp. 1–6, 2018. doi: 10.1016/j.coviro.2018.06.001
-
[44]
W. McCormick and L. A. Mermel, “The basic reproductive number and particle-to-plaque ratio: Comparison of these two parameters of viral infectivity,”Virology Journal, vol. 18, article 92, 2021. doi: 10.1186/s12985-021-01566-4
-
[45]
Single-virus tracking: From imaging methodologies to virological applications,
S.-L. Liu, Z.-G. Wang, H.-Y. Xie, A. A. Liu, D. C. Lamb, and D.-W. Pang, “Single-virus tracking: From imaging methodologies to virological applications,”Chemical Reviews, vol. 120, no. 3, pp. 1936–1979, 2020. doi: 10.1021/acs.chemrev.9b00692
-
[46]
Imaging, tracking and computa- tional analyses of virus entry and egress with the cytoskeleton,
I.-H. Wang, C. J. Burckhardt, A. Yakimovich, and U. F. Greber, “Imaging, tracking and computa- tional analyses of virus entry and egress with the cytoskeleton,”Viruses, vol. 10, no. 4, article 166,
-
[47]
doi: 10.3390/v10040166
-
[48]
L. Nathan and S. Daniel, “Single virion tracking microscopy for the study of virus entry processes in live cells and biomimetic platforms,” inPhysical Virology: Virus Structure and Mechanics, U. F. Greber, ed.,Advances in Experimental Medicine and Biology, vol. 1215, pp. 13–43, Cham: Springer, 2019. doi:10.1007/978-3-030-14741-9_2
-
[49]
On the mathematical foundations of theoretical statistics,
R. A. Fisher, “On the mathematical foundations of theoretical statistics,”Philosophical Transactions of the Royal Society of London. Series A, vol. 222, pp. 309–368, 1922. doi: 10.1098/rsta.1922.0009
-
[50]
Information and the accuracy attainable in the estimation of statistical parameters,
C. R. Rao, “Information and the accuracy attainable in the estimation of statistical parameters,” Bulletin of the Calcutta Mathematical Society, vol. 37, no. 3, pp. 81–91, 1945
1945
-
[51]
Cramér,Mathematical Methods of Statistics
H. Cramér,Mathematical Methods of Statistics. Princeton, NJ: Princeton University Press, 1946
1946
-
[52]
I. J. Hirst, W. J. R. Thomas, R. A. Davies, and S. P. Muench, “CryoEM grid preparation: A closer look at advancements and impact of preparation mode and new approaches,”Biochemical Society Transactions, vol. 52, no. 3, pp. 1529–1537, 2024. doi: 10.1042/BST20231553
-
[53]
Addressing preferred specimen orientation in single-particle cryo-EM through tilting,
Y. Z. Tan, P. R. Baldwin, J. H. Davis, J. R. Williamson, C. S. Potter, B. Carragher, and D. Lyumkis, “Addressing preferred specimen orientation in single-particle cryo-EM through tilting,”Nature Methods, vol. 14, pp. 793–796, 2017. doi: 10.1038/nmeth.4347. 173
-
[54]
S. Wiedemann and R. Heckel, “A deep learning method for simultaneous denoising and miss- ing wedge reconstruction in cryogenic electron tomography,”Nature Communications, vol. 15, article 8255, 2024. doi: 10.1038/s41467-024-51438-y
-
[55]
Imaging and manipulation of single viruses by atomic force microscopy,
M. Baclayon, G. J. L. Wuite, and W. H. Roos, “Imaging and manipulation of single viruses by atomic force microscopy,”Soft Matter, vol. 6, no. 21, pp. 5273–5285, 2010. doi: 10.1039/B923992H
-
[56]
The motion and precipitation of suspensoids in divergent electric fields,
H. A. Pohl, “The motion and precipitation of suspensoids in divergent electric fields,”Journal of Applied Physics, vol. 22, no. 7, pp. 869–871, 1951. doi: 10.1063/1.1700065
-
[57]
Need for speed: Examining protein behavior during cryoEM grid preparation at different timescales,
D. P. Klebl, M. S. C. Gravett, D. Kontziampasis, D. J. Wright, R. S. Bon, D. C. F. Monteiro, M. Trebbin, F. Sobott, H. D. White, M. C. Darrow, R. F. Thompson, and S. P. Muench, “Need for speed: Examining protein behavior during cryoEM grid preparation at different timescales,” Structure, vol. 28, no. 11, pp. 1238–1248.e4, 2020. doi: 10.1016/j.str.2020.07.018
-
[58]
Time to revisit the endpoint dilution assay and to replace the TCID50 as a measure of a virus sample’s infection concentration,
D. Cresta, D. C. Warren, C. Quirouette, A. P. Smith, L. C. Lane, A. M. Smith, and C. A. A. Beauchemin, “Time to revisit the endpoint dilution assay and to replace the TCID50 as a measure of a virus sample’s infection concentration,”PLOS Computational Biology, vol. 17, no. 10, e1009480,
-
[59]
doi: 10.1371/journal.pcbi.1009480
-
[60]
D. Cresta, D. C. Warren, C. Quirouette, A. P. Smith, L. C. Lane, A. M. Smith, and C. A. A. Beauchemin, “Correction: Time to revisit the endpoint dilution assay and to replace the TCID50 as a measure of a virus sample’s infection concentration,”PLOS Computational Biology, vol. 19, no. 1, e1010877, 2023. doi: 10.1371/journal.pcbi.1010877
-
[61]
On the calculation of TCID50 for quantitation of virus infectivity,
C. Lei, J. Yang, J. Hu, and X. Sun, “On the calculation of TCID50 for quantitation of virus infectivity,”Virologica Sinica, vol. 36, no. 1, pp. 141–144, 2021. doi: 10.1007/s12250-020-00230-5
-
[62]
Inverse problems: A Bayesian perspective,
A. M. Stuart, “Inverse problems: A Bayesian perspective,”Acta Numerica, vol. 19, pp. 451–559,
-
[63]
doi: 10.1017/S0962492910000061
-
[64]
Y.-C. Hsieh, M. Delarue, H. Orland, and P. Koehl, “Analyzing the geometry and dynamics of viral structures: A review of computational approaches based on alpha shape theory, normal mode analysis, and Poisson–Boltzmann theories,”Viruses, vol. 15, no. 6, article 1366, 2023. doi: 10.3390/v15061366
-
[65]
Physical properties of the HIV-1 capsid from all-atom molecular dynamics simulations,
J. R. Perilla and K. Schulten, “Physical properties of the HIV-1 capsid from all-atom molecular dynamics simulations,”Nature Communications, vol. 8, article 15959, 2017. doi: 10.1038/ncomms15959
-
[66]
R. F. Bruinsma, G. J. L. Wuite, and W. H. Roos, “Physics of viral dynamics,”Nature Reviews Physics, vol. 3, no. 2, pp. 76–91, 2021. doi: 10.1038/s42254-020-00267-1
-
[67]
Two detailed plaque assay protocols for the quantification of infectious SARS-CoV-2,
E. J. Mendoza, K. Manguiat, H. Wood, and M. Drebot, “Two detailed plaque assay protocols for the quantification of infectious SARS-CoV-2,”Current Protocols in Microbiology, vol. 57, no. 1, e105, 2020. doi: 10.1002/cpmc.105. 174
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.