Multiplicative modulations in hue-selective cells enhance unique hue representation
Pith reviewed 2026-05-25 09:20 UTC · model grok-4.3
The pith
Multiplicative modulations between color channels allow model V4 neurons to encode unique hues.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
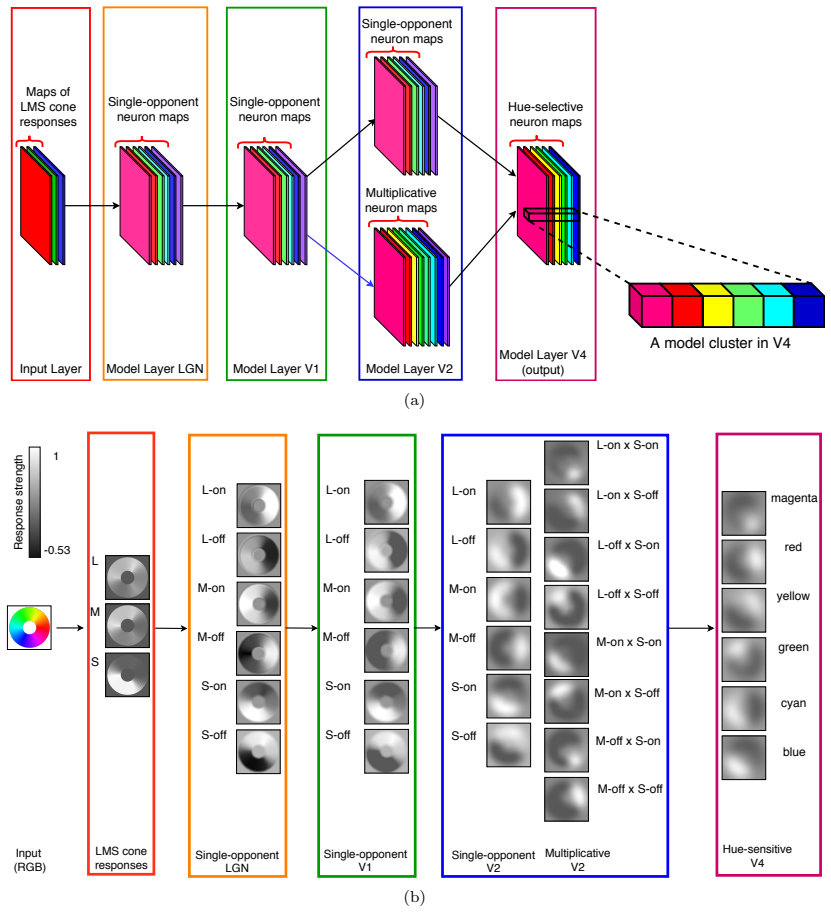
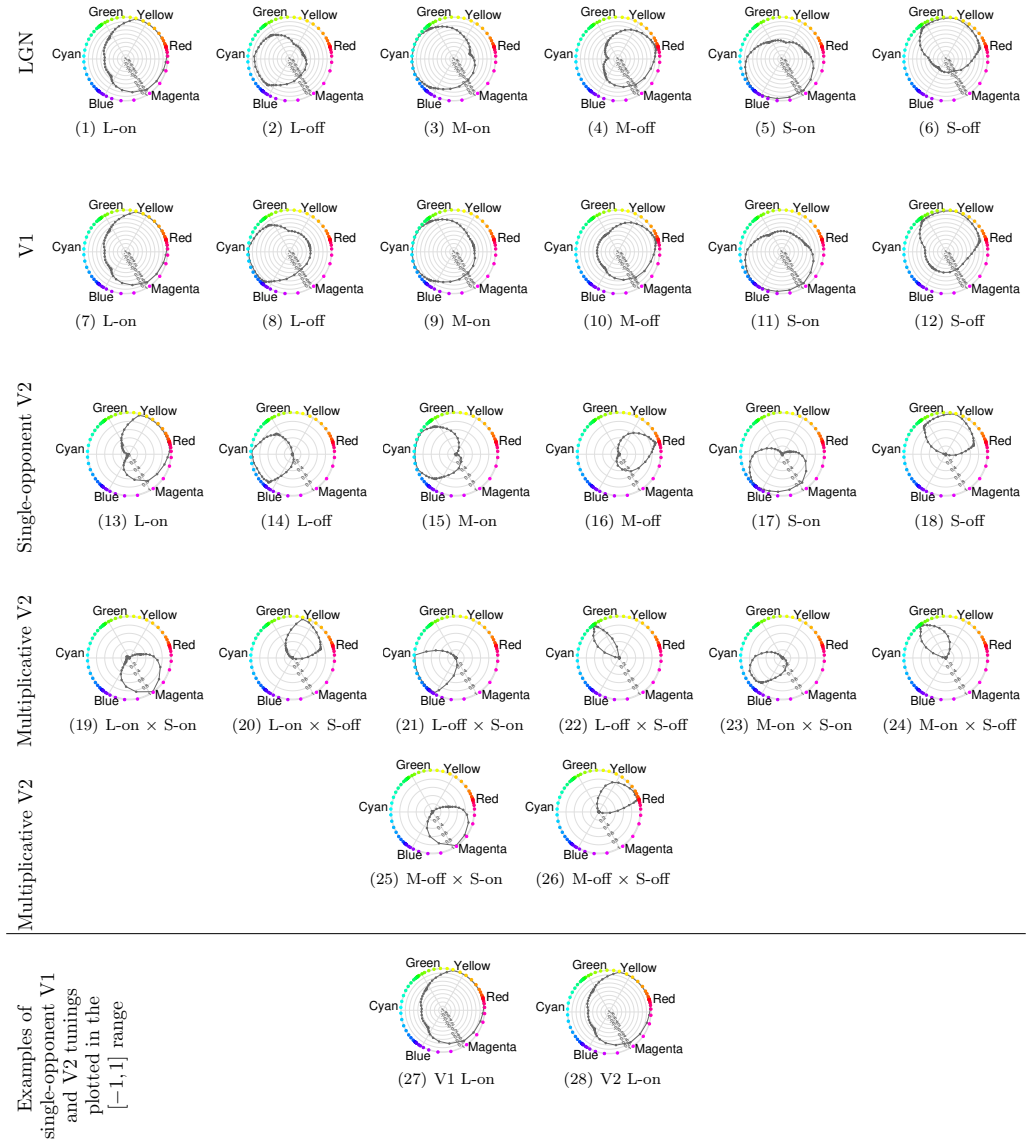
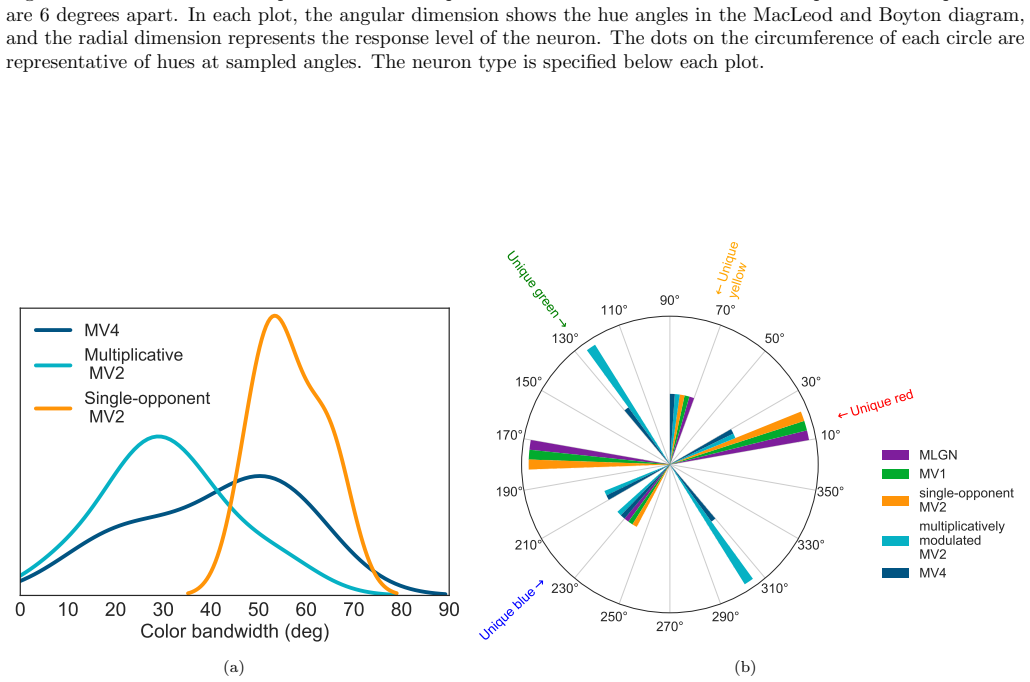
The authors show through simulation that multiplicative modulations applied to cone-opponent signals, when embedded in a layered network inspired by cortical processing, enable neurons at the V4 stage to develop tuning for unique hues while also improving representation of intermediate chromatic directions.
What carries the argument
Multiplicative modulations between opponent channels, introduced as an additional nonlinearity after initial rectifications in a hierarchical network.
If this is right
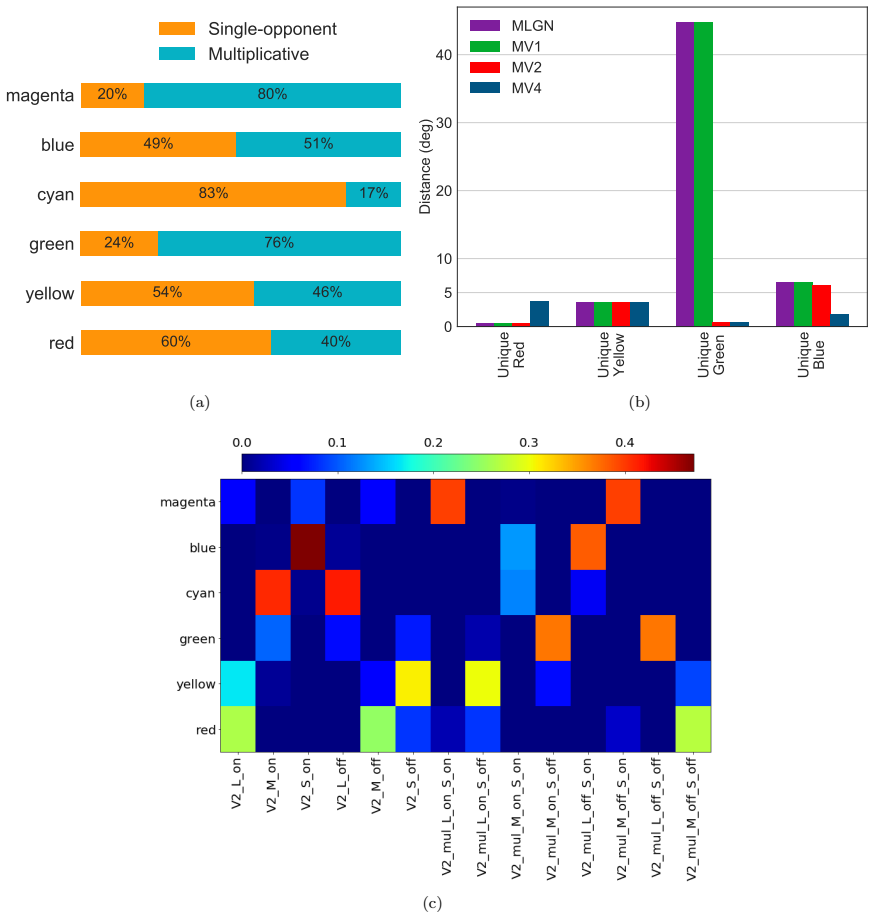
- Multiplicative modulations contribute substantially to encoding hues along intermediate directions in the MacLeod-Boynton diagram.
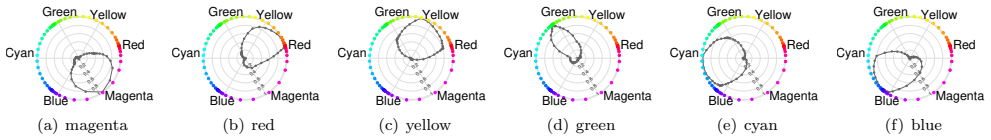
- Neurons in a model of area V4 acquire the capacity to represent the four unique hues.
- The model's neuron responses reproduce patterns seen in biological hue-selective cells.
- Incremental addition of nonlinearities from cone input produces the observed transformation to perceptual hue representation.
Where Pith is reading between the lines
- The same modulation rule could be tested for its effect on hue constancy under changing illumination.
- Recording from V4 while pharmacologically altering candidate multiplicative interactions would provide a direct test of the mechanism.
- The layered structure supplies a template for predicting how hue tuning evolves across earlier visual areas.
Load-bearing premise
The particular form and locations chosen for the multiplicative modulations correspond to the nonlinear operations actually performed along the primate color pathway.
What would settle it
Removing the multiplicative terms from the model and observing that intermediate-hue responses in the V4 layer remain essentially unchanged would falsify the claim of their significant contribution.
Figures
read the original abstract
There is still much to understand about the color processing mechanisms in the brain and the transformation from cone-opponent representations to perceptual hues. Moreover, it is unclear which areas(s) in the brain represent unique hues. We propose a hierarchical model inspired by the neuronal mechanisms in the brain for local hue representation, which reveals the contributions of each visual cortical area in hue representation. Local hue encoding is achieved through incrementally increasing processing nonlinearities beginning with cone input. Besides employing nonlinear rectifications, we propose multiplicative modulations as a form of nonlinearity. Our simulation results indicate that multiplicative modulations have significant contributions in encoding of hues along intermediate directions in the MacLeod-Boynton diagram and that model V4 neurons have the capacity to encode unique hues. Additionally, responses of our model neurons resemble those of biological color cells, suggesting that our model provides a novel formulation of the brain's color processing pathway.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes a hierarchical computational model of hue representation in the primate visual pathway, starting from cone inputs and incrementally adding nonlinear rectifications followed by multiplicative modulations between channels. Simulations are used to argue that these multiplicative modulations make significant contributions to encoding hues along intermediate directions in the MacLeod-Boynton diagram and that the V4 stage possesses the capacity to represent unique hues, with model neuron responses resembling biological color cells.
Significance. If the modeling choices are shown to be physiologically grounded and the simulation outcomes are quantitatively validated, the work would provide a concrete, bottom-up account of how unique-hue selectivity can emerge in V4 and would highlight multiplicative interactions as a distinct computational motif in cortical color processing. The absence of circularity (model is constructed from cone inputs rather than fitted to the target data) is a positive feature.
major comments (3)
- [Abstract] Abstract: the central claim that 'multiplicative modulations have significant contributions' rests on simulation outcomes that supply neither quantitative error bars, statistical tests, nor any measure of effect size; without these the reported resemblance to biological cells cannot be evaluated.
- [Results] Simulation results (implicit in abstract and results narrative): no comparison is presented against alternative nonlinearities (e.g., different rectification schemes or additive interactions) or against an ablation that removes the multiplicative stage, so it is impossible to determine whether the reported improvement in intermediate-hue encoding is attributable to the specific modulation rule or to other model components.
- [Methods] Methods / model description: the specific placement and functional form of the multiplicative modulations are stated to be 'inspired by neuronal mechanisms' but no independent physiological constraint, parameter count, or cross-validation against recorded V4 responses is supplied; this leaves the claimed biological relevance of the modulation rule untested.
minor comments (1)
- [Methods] The number of free parameters adjusted to produce the reported cell-like responses is never stated, making reproducibility and assessment of model complexity difficult.
Simulated Author's Rebuttal
We thank the referee for the constructive comments, which highlight areas where the manuscript can be strengthened. We address each major comment below and indicate the revisions we will make.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central claim that 'multiplicative modulations have significant contributions' rests on simulation outcomes that supply neither quantitative error bars, statistical tests, nor any measure of effect size; without these the reported resemblance to biological cells cannot be evaluated.
Authors: We agree that quantitative support is needed to substantiate the claims. In the revised manuscript we will report means and standard deviations across multiple simulation runs (with error bars), compute effect sizes for the contribution of multiplicative modulations, and apply appropriate statistical tests to the comparisons with biological cell responses. revision: yes
-
Referee: [Results] Simulation results (implicit in abstract and results narrative): no comparison is presented against alternative nonlinearities (e.g., different rectification schemes or additive interactions) or against an ablation that removes the multiplicative stage, so it is impossible to determine whether the reported improvement in intermediate-hue encoding is attributable to the specific modulation rule or to other model components.
Authors: The model is constructed incrementally to isolate the effect of each added nonlinearity. To directly address the concern we will include an ablation analysis that removes the multiplicative stage and additional control simulations using only additive combinations or alternative rectification functions, allowing quantitative attribution of the observed improvements. revision: yes
-
Referee: [Methods] Methods / model description: the specific placement and functional form of the multiplicative modulations are stated to be 'inspired by neuronal mechanisms' but no independent physiological constraint, parameter count, or cross-validation against recorded V4 responses is supplied; this leaves the claimed biological relevance of the modulation rule untested.
Authors: The placement and form draw from established cortical mechanisms (e.g., cross-orientation suppression and divisive normalization) cited in the introduction and methods. We will expand the methods section to list the exact parameter count, provide explicit citations for each modeling choice, and explicitly state that the model is not fitted to V4 data. Direct cross-validation against recorded V4 responses is outside the present bottom-up scope but will be noted as a limitation and direction for future work. revision: partial
Circularity Check
No significant circularity; model outputs are simulation results from bottom-up construction
full rationale
The paper builds a hierarchical model starting from cone inputs, incrementally adding rectifications and a proposed multiplicative modulation rule described as inspired by biology. Simulation results on hue encoding and unique-hue capacity in the V4 stage are direct outputs of running this model; no equation or result is shown to reduce by construction to a fitted parameter, self-citation, or input data that defines the target claim. The modulation rule is an explicit modeling choice rather than a derived necessity, but this does not create circularity under the defined patterns because the reported contributions are not tautological with the inputs.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The primate color pathway can be approximated by a feed-forward hierarchy of areas V1-V2-V4 with successively stronger nonlinearities.
Reference graph
Works this paper leans on
-
[1]
Space and time maps of cone photoreceptor signals in macaque lateral geniculate nucleus,
R. C. Reid and R. M. Shapley, “Space and time maps of cone photoreceptor signals in macaque lateral geniculate nucleus,” Journal of Neuroscience, vol. 22, no. 14, pp. 6158–6175, 2002. [Online]. Available: http://www.jneurosci.org/content/22/14/6158
work page 2002
-
[2]
Outlines of a theory of the light sense
E. Hering, “Outlines of a theory of the light sense.” 1964. 10 Table 1: The choice of weights for model V4 cells used for hue reconstruction in a few example hues. Groundtruth hue (deg) Model V4 neuron Red Yellow Green Cyan Blue Magenta red (360) 0.9903 0.0034 0.0006 0.0007 0.0006 0.0014 Yellow (60) 0.0010 0.9950 0.0009 0.0021 0.0004 0.0005 lavender (270)...
work page 1964
-
[3]
Chromatic mechanisms in lateral geniculate nucleus of macaque
A. M. Derrington, J. Krauskopf, and P. Lennie, “Chromatic mechanisms in lateral geniculate nucleus of macaque.” The Journal of physiology , vol. 357, no. 1, pp. 241–265, 1984
work page 1984
-
[4]
Some transformations of color information from lateral geniculate nucleus to striate cortex,
R. L. De Valois, N. P. Cottaris, S. D. Elfar, L. E. Mahon, and J. A. Wilson, “Some transformations of color information from lateral geniculate nucleus to striate cortex,” Proceedings of the National Academy of Sciences, vol. 97, no. 9, pp. 4997–5002, 2000
work page 2000
-
[5]
Specificity of cone inputs to macaque retinal gan- glion cells,
H. Sun, H. E. Smithson, Q. Zaidi, and B. B. Lee, “Specificity of cone inputs to macaque retinal gan- glion cells,” Journal of neurophysiology, vol. 95, no. 2, pp. 837–849, 2006
work page 2006
-
[6]
Chromaticity di- agram showing cone excitation by stimuli of equal lu- minance,
D. I. MacLeod and R. M. Boynton, “Chromaticity di- agram showing cone excitation by stimuli of equal lu- minance,” JOSA, vol. 69, no. 8, pp. 1183–1186, 1979
work page 1979
-
[7]
Neural selectivity for hue and saturation of colour in the pri- mary visual cortex of the monkey,
A. Hanazawa, H. Komatsu, and I. Murakami, “Neural selectivity for hue and saturation of colour in the pri- mary visual cortex of the monkey,” European Journal of Neuroscience, vol. 12, no. 5, pp. 1753–1763, 2000
work page 2000
-
[8]
Hue selectivity in human visual cortex revealed by functional magnetic resonance imaging,
I. Kuriki, P. Sun, K. Ueno, K. Tanaka, and K. Cheng, “Hue selectivity in human visual cortex revealed by functional magnetic resonance imaging,” Cerebral Cortex, vol. 25, no. 12, pp. 4869–4884, 2015
work page 2015
-
[9]
Cone inputs in macaque primary visual cortex,
E. N. Johnson, M. J. Hawken, and R. Shapley, “Cone inputs in macaque primary visual cortex,” Journal of Neurophysiology, vol. 91, no. 6, pp. 2501–2514, 2004
work page 2004
-
[10]
Neural mechanisms for color perception in the primary visual cortex,
R. Shapley and M. Hawken, “Neural mechanisms for color perception in the primary visual cortex,” Cur- rent opinion in neurobiology , vol. 12, no. 4, pp. 426– 432, 2002
work page 2002
-
[11]
Chromatic mechanisms in striate cortex of macaque,
P. Lennie, J. Krauskopf, and G. Sclar, “Chromatic mechanisms in striate cortex of macaque,” Journal of Neuroscience, vol. 10, no. 2, pp. 649–669, 1990
work page 1990
-
[12]
Representation of color stimuli in awake macaque primary visual cortex,
T. Wachtler, T. J. Sejnowski, and T. D. Albright, “Representation of color stimuli in awake macaque primary visual cortex,” Neuron, vol. 37, no. 4, pp. 681–691, 2003
work page 2003
-
[13]
T. Namima, M. Yasuda, T. Banno, G. Okazawa, and H. Komatsu, “Effects of luminance contrast on the color selectivity of neurons in the macaque area v4 and inferior temporal cortex,” Journal of Neuro- science, vol. 34, no. 45, pp. 14 934–14 947, 2014
work page 2014
-
[14]
Special- ized color modules in macaque extrastriate cortex,
B. R. Conway, S. Moeller, and D. Y. Tsao, “Special- ized color modules in macaque extrastriate cortex,” Neuron, vol. 56, no. 3, pp. 560–573, 2007
work page 2007
-
[15]
Percep- tual color map in macaque visual area v4,
M. Li, F. Liu, M. Juusola, and S. Tang, “Percep- tual color map in macaque visual area v4,” Journal of Neuroscience, vol. 34, no. 1, pp. 202–217, 2014
work page 2014
-
[16]
B. R. Conway and D. Y. Tsao, “Color-tuned neu- rons are spatially clustered according to color prefer- ence within alert macaque posterior inferior temporal cortex,” Proceedings of the National Academy of Sci- ences, vol. 106, no. 42, pp. 18 034–18 039, 2009
work page 2009
-
[17]
Why have multiple cortical areas?
H. B. Barlow, “Why have multiple cortical areas?” Vision research, vol. 26, no. 1, pp. 81–90, 1986
work page 1986
-
[18]
A spatially organized representation of colour in macaque corti- cal area v2,
Y. Xiao, Y. Wang, and D. J. Felleman, “A spatially organized representation of colour in macaque corti- cal area v2,” Nature, vol. 421, no. 6922, pp. 535–539, 2003
work page 2003
-
[19]
K. S. Bohon, K. L. Hermann, T. Hansen, and B. R. Conway, “Representation of perceptual color space in macaque posterior inferior temporal cortex (the v4 complex),” eNeuro, vol. 3, no. 4, pp. ENEURO–0039, 2016
work page 2016
-
[20]
Spectral properties of v4 neurons in the macaque,
S. J. Schein and R. Desimone, “Spectral properties of v4 neurons in the macaque,”Journal of Neuroscience, vol. 10, no. 10, pp. 3369–3389, 1990
work page 1990
-
[21]
The representation of colours in the cerebral cortex,
S. Zeki, “The representation of colours in the cerebral cortex,” Nature, vol. 284, no. 5755, pp. 412–418, 1980
work page 1980
-
[22]
Variations in normal color vision. ii. unique hues,
M. A. Webster, E. Miyahara, G. Malkoc, and V. E. Raker, “Variations in normal color vision. ii. unique hues,” JOSA A , vol. 17, no. 9, pp. 1545–1555, 2000
work page 2000
-
[23]
The cone inputs to the unique-hue mechanisms,
S. M. Wuerger, P. Atkinson, and S. Cropper, “The cone inputs to the unique-hue mechanisms,” Vision research, vol. 45, no. 25, pp. 3210–3223, 2005
work page 2005
-
[24]
C. M. Stoughton and B. R. Conway, “Neural basis for unique hues,” Current Biology , vol. 18, no. 16, pp. R698–R699, 2008. 11
work page 2008
-
[25]
Color selectivity of neurons in the inferior tempo- ral cortex of the awake macaque monkey,
H. Komatsu, Y. Ideura, S. Kaji, and S. Yamane, “Color selectivity of neurons in the inferior tempo- ral cortex of the awake macaque monkey,” Journal of Neuroscience, vol. 12, no. 2, pp. 408–424, 1992
work page 1992
-
[26]
A neural basis for unique hues?
J. Mollon, “A neural basis for unique hues?” Current Biology, vol. 19, no. 11, pp. R441–R442, 2009
work page 2009
-
[27]
Q. Zaidi, R. Ennis, L. Wool, S. Komban, J.-M. Alonso, B. Conway, G. Gagin, and K. Bohon, “The enigma of unique hues,” in I-PERCEPTION, vol. 5, no. 4. PION LTD 207 BRONDESBURY PARK, LONDON NW2 5JN, ENGLAND, 2014, pp. 421–421
work page 2014
-
[28]
Color categoriza- tion and color constancy in a neural network model of v4,
P. A. Dufort and C. J. Lumsden, “Color categoriza- tion and color constancy in a neural network model of v4,” Biological cybernetics, vol. 65, no. 4, pp. 293– 303, 1991
work page 1991
-
[29]
A multistage neural network for color constancy and color induction,
S. M. Courtney, L. H. Finkel, and G. Buchsbaum, “A multistage neural network for color constancy and color induction,” IEEE Transactions on Neural Net- works, vol. 6, no. 4, pp. 972–985, 1995
work page 1995
-
[30]
A model of color vision based on cortical reentry,
J. Wray and G. M. Edelman, “A model of color vision based on cortical reentry,” Cerebral Cortex , vol. 6, no. 5, pp. 701–716, 1996
work page 1996
-
[31]
R. L. De Valois and K. K. De Valois, “A multi-stage color model,” Vision research , vol. 33, no. 8, pp. 1053–1065, 1993
work page 1993
-
[32]
Seeing white: Qualia in the context of decoding population codes,
S. R. Lehky and T. J. Sejnowski, “Seeing white: Qualia in the context of decoding population codes,” Neural computation , vol. 11, no. 6, pp. 1261–1280, 1999
work page 1999
-
[33]
The tale of the neuroscientists and the computer: why mechanistic theory matters,
J. W. Brown, “The tale of the neuroscientists and the computer: why mechanistic theory matters,” Fron- tiers in neuroscience , vol. 8, 2014
work page 2014
-
[34]
E. Miyahara, “Focal colors and unique hues,” Percep- tual and motor skills , vol. 97, no. 3 suppl, pp. 1038– 1042, 2003
work page 2003
-
[35]
Chromatic properties of neurons in macaque area v2,
D. C. Kiper, S. B. Fenstemaker, and K. R. Gegenfurt- ner, “Chromatic properties of neurons in macaque area v2,” Visual neuroscience , vol. 14, no. 6, pp. 1061–1072, 1997
work page 1997
-
[36]
An interior-point method for large-scale l1-regularized logistic regres- sion,
K. Koh, S.-J. Kim, and S. Boyd, “An interior-point method for large-scale l1-regularized logistic regres- sion,” Journal of Machine learning research , vol. 8, no. Jul, pp. 1519–1555, 2007
work page 2007
-
[37]
Converting rgb images to lms cone activations,
J. B. D. Paula, “Converting rgb images to lms cone activations,” Department of Computer Sciences, The University of Texas at Austin, Tech. Rep., 2006, technical Report 06-49. [Online]. Available: http://nn.cs.utexas.edu/?depaula:utcstr06-49
work page 2006
-
[38]
A. L. Rothenstein, A. Zaharescu, and J. K. Tsotsos, TarzaNN : A General Purpose Neural Network Simu- lator for Visual Attention Modeling . Berlin, Heidel- berg: Springer Berlin Heidelberg, 2005, pp. 159–167
work page 2005
-
[39]
Metamers of the ventral stream,
J. Freeman and E. P. Simoncelli, “Metamers of the ventral stream,” Nature neuroscience, vol. 14, no. 9, pp. 1195–1201, 2011
work page 2011
-
[40]
Color in the cortex: single-and double-opponent cells,
R. Shapley and M. J. Hawken, “Color in the cortex: single-and double-opponent cells,” Vision research , vol. 51, no. 7, pp. 701–717, 2011
work page 2011
-
[41]
Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (v- 1),
B. R. Conway, “Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (v- 1),” Journal of Neuroscience, vol. 21, no. 8, pp. 2768– 2783, 2001
work page 2001
-
[42]
Network dissection: Quantifying inter- pretability of deep visual representations,
D. Bau, B. Zhou, A. Khosla, A. Oliva, and A. Torralba, “Network dissection: Quantifying inter- pretability of deep visual representations,” in Pro- ceedings of the IEEE Conference on Computer Vision and Pattern Recognition, 2017, pp. 6541–6549
work page 2017
-
[43]
The build- ing blocks of interpretability,
C. Olah, A. Satyanarayan, I. Johnson, S. Carter, L. Schubert, K. Ye, and A. Mordvintsev, “The build- ing blocks of interpretability,” Distill, vol. 3, no. 3, p. e10, 2018. Acknowledgements This research was supported by several sources for which the authors are grateful: Air Force Office of Scientific Re- search (FA9550-18-1-0054), the Canada Research Chairs...
work page 2018
-
[44]
In doing so, we designed each neuron in our network according to the existing findings of the brain
Our goal was to introduce a biologically inspired model that would help in understanding hue encod- ing mechanisms in the brain. In doing so, we designed each neuron in our network according to the existing findings of the brain. For example, the receptive field profile and the weights from cones to single-opponent cells in our model LGN layer were set based...
-
[45]
Often, the learned features in the first hidden layer are compared with biological V1 neurons
One challenge in convolutional neural networks is in- terpreting the learned features in the hidden layers. Often, the learned features in the first hidden layer are compared with biological V1 neurons. However, learned features in deeper layers are difficult to ex- plain. There have been attempts to understand and interpret hidden layer features [42, 43]. H...
-
[46]
Nonetheless, even with such data acces- sible to us, we would not have been able to use a CNN model
In this work, we did not have access to any cell record- ing data. Nonetheless, even with such data acces- sible to us, we would not have been able to use a CNN model. Often, cell recording data is limited and sparse and not enough for learning the massive num- ber of parameters in a CNN. We acknowledge that a certain set of parameters in our model were s...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.