Moving backward to go faster: Diatom-inspired sliding reveals efficient modes of locomotion
Pith reviewed 2026-06-27 11:43 UTC · model grok-4.3
The pith
Sliding between stacked cells propels chains faster and more efficiently than undulatory bending.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Sliding between stacked elongated cells generates internal shear that drives propulsion opposite to classical undulatory swimming, while achieving higher speeds and greater energetic efficiency. Optimal performance occurs at wavelengths much larger than the chain length and at cell aspect ratios consistent with those observed in natural diatom colonies.
What carries the argument
Internal shear generated by sliding between neighboring cells in a hydrodynamic chain model of diatom colonies.
If this is right
- Propulsion direction is opposite to the traveling wave, unlike classical undulatory gaits.
- Speeds exceed those achieved by bending-based swimmers under comparable conditions.
- Energetic efficiency is higher than in undulatory locomotion.
- Performance peaks at wavelengths much larger than chain length and at specific cell aspect ratios.
Where Pith is reading between the lines
- The sliding mechanism may operate in other biological chain assemblies beyond diatoms.
- It supplies concrete design rules for building efficient sliding-based microswimmers or robotic chains.
- The match between optimal aspect ratios and natural diatom colonies supports hydrodynamic efficiency as an evolutionary driver.
Load-bearing premise
Hydrodynamic interactions in the chain are dominated by sliding shear without major contributions from cell flexibility or external flow variations.
What would settle it
A simulation or physical experiment in which preventing sliding or shortening wavelengths to match chain length fails to reduce speed or efficiency relative to undulatory bending would falsify the central claim.
Figures
read the original abstract
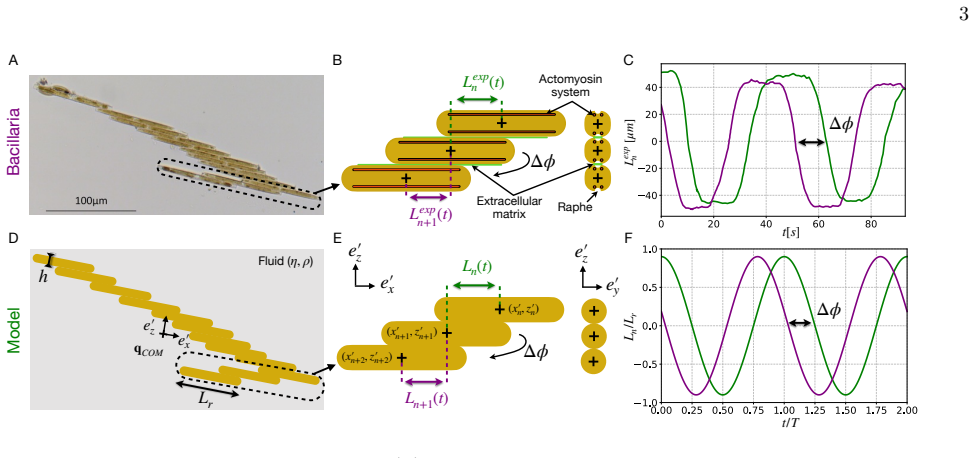
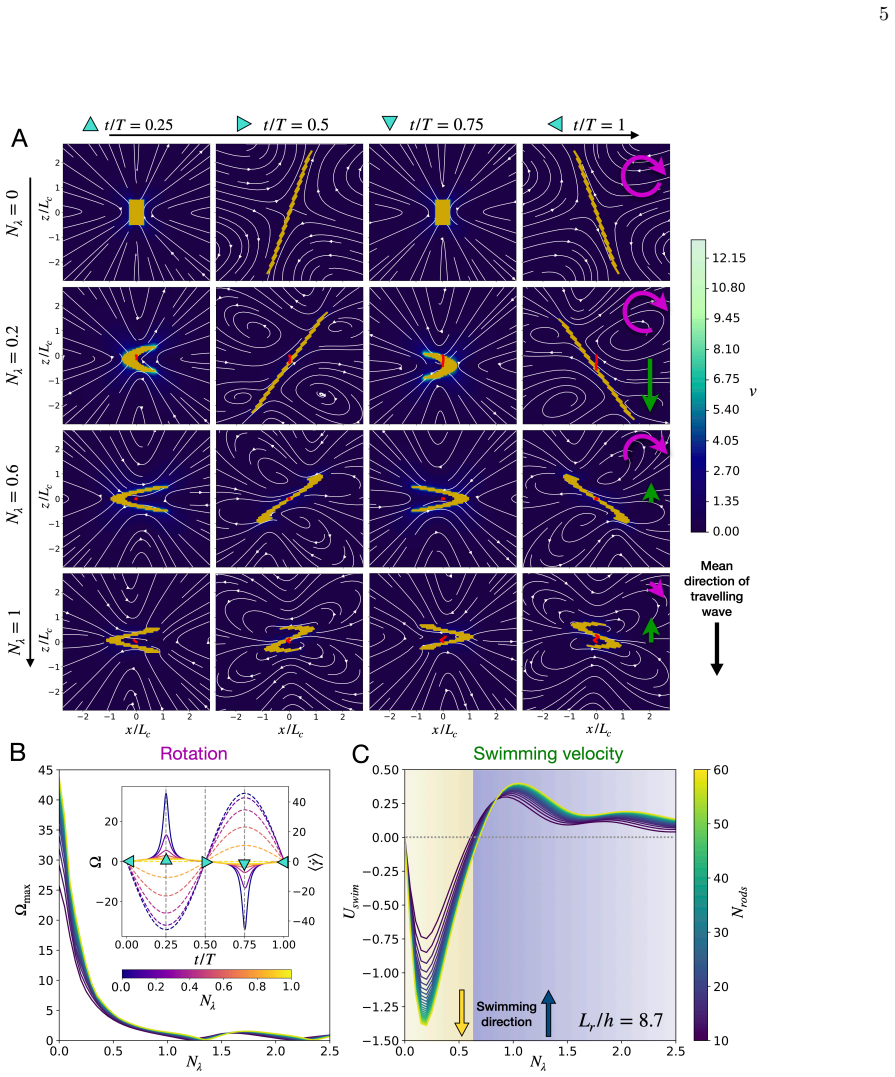
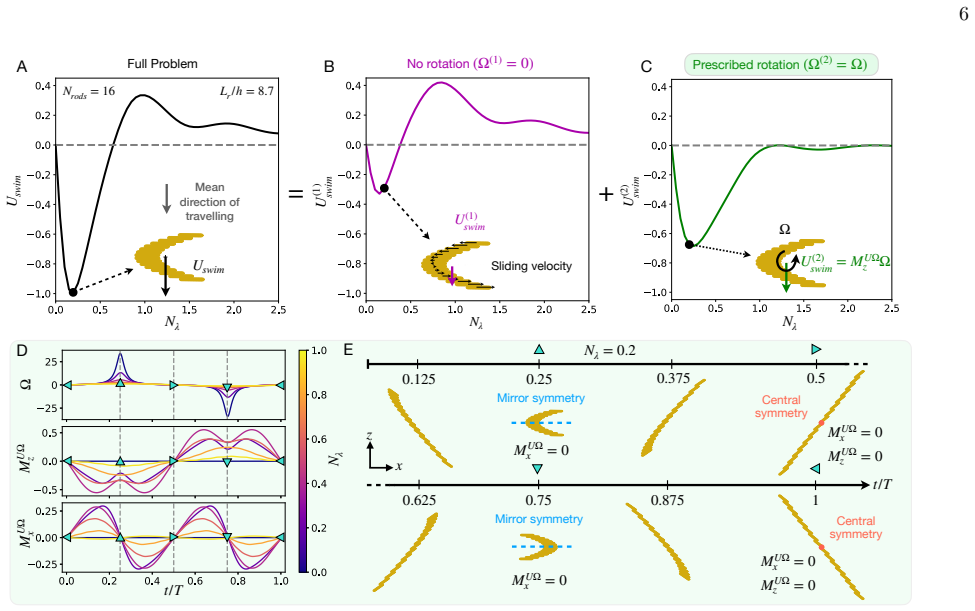
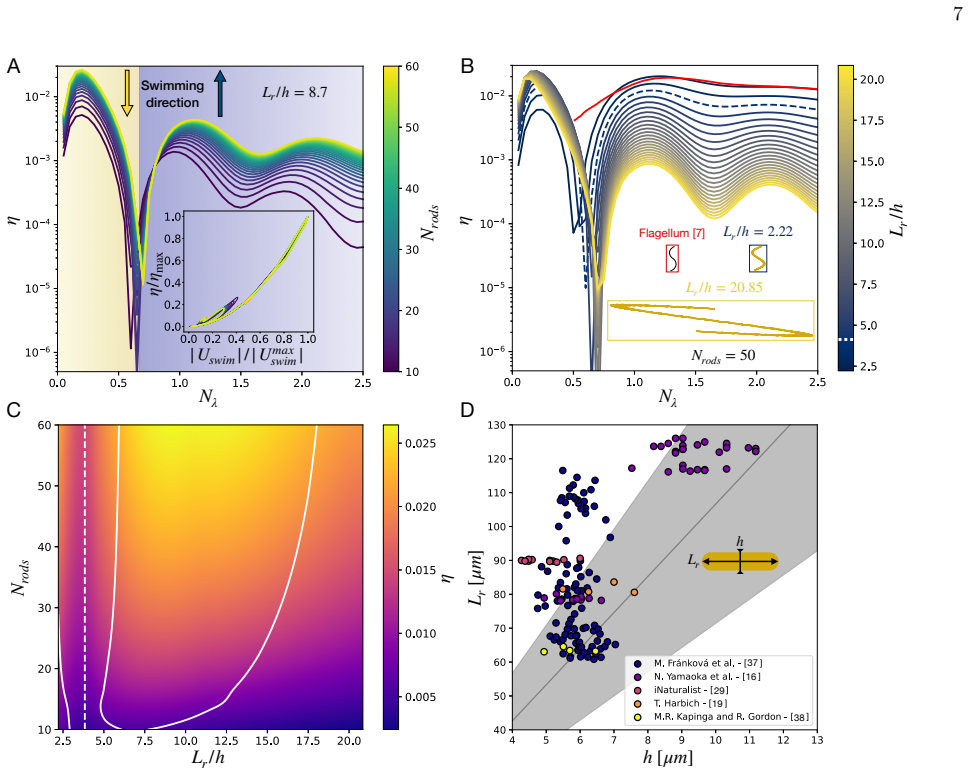
Across biological scales, from sperm cells to whales, locomotion commonly relies on undulatory gaits, in which traveling deformation waves interact with the surrounding fluid to generate thrust opposite to the direction of wave propagation. In viscous environments, microorganism locomotion is classically understood in terms of undulatory bending of slender filaments such as flagella, with optimal propulsion achieved when the deformation wavelength is comparable to the swimmer length. Inspired by diatom colonies, we identify a fundamentally different swimming mechanism based on sliding between neighboring elements within a chain. We show that sliding between stacked elongated cells generates internal shear that drives propulsion opposite to classical undulatory swimming, while achieving higher speeds and greater energetic efficiency. Remarkably, optimal performance occurs at wavelengths much larger than the chain length and at cell aspect ratios consistent with those observed in natural diatom colonies, suggesting that hydrodynamic efficiency may constitute an evolutionary selective pressure in diatom chains. Together, these results identify sliding as a previously overlooked mode of locomotion in multicellular assemblies and suggest new design principles for efficient bio-inspired microswimmers and swarm robotic systems.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that sliding between stacked elongated cells in diatom-inspired chains generates internal shear driving propulsion opposite to classical undulatory swimming, while achieving higher speeds and greater energetic efficiency. Optimal performance occurs at wavelengths much larger than the chain length and at cell aspect ratios matching natural diatom colonies, suggesting hydrodynamic efficiency as an evolutionary pressure and new design principles for microswimmers.
Significance. If the hydrodynamic model holds, this identifies sliding as a previously overlooked efficient locomotion mode in multicellular assemblies, potentially explaining observed diatom colony structures and offering alternatives to undulatory propulsion in viscous media. The parameter-free aspects of the optima and biological consistency add value if quantitatively validated.
major comments (1)
- [Hydrodynamic model] Model and results sections: the central efficiency and directionality claims rest on the assumption that sliding shear dominates hydrodynamic interactions; without explicit comparison or sensitivity analysis to unmodeled effects such as cell flexibility or external flow variations (as flagged in the weakest assumption), the ordering of speeds and efficiency versus undulatory modes cannot be confirmed as robust.
minor comments (2)
- [Abstract] Abstract: strong quantitative claims (higher speeds, greater efficiency) are made without any reported numbers, error bars, or key parameter values; adding one or two representative results would improve clarity.
- Notation: define all symbols (e.g., wavelength, aspect ratio, efficiency metric) at first use and ensure consistency between text and any figures.
Simulated Author's Rebuttal
We thank the referee for their constructive comments. We address the single major comment below.
read point-by-point responses
-
Referee: [Hydrodynamic model] Model and results sections: the central efficiency and directionality claims rest on the assumption that sliding shear dominates hydrodynamic interactions; without explicit comparison or sensitivity analysis to unmodeled effects such as cell flexibility or external flow variations (as flagged in the weakest assumption), the ordering of speeds and efficiency versus undulatory modes cannot be confirmed as robust.
Authors: We agree that the central claims depend on sliding shear dominating the hydrodynamics and that the manuscript flags cell flexibility and external flow variations as the weakest assumptions. The current model isolates the sliding mechanism under rigid-cell, no-external-flow conditions at low Reynolds number, but does not include explicit sensitivity tests. In revision we will add a dedicated sensitivity subsection that perturbs cell rigidity (via a low-stiffness beam model) and superimposes weak uniform external flows. We will show that the reported directionality (propulsion opposite undulatory swimming) and efficiency ordering remain qualitatively unchanged within the biologically relevant parameter range. This addition will directly address the robustness concern. revision: yes
Circularity Check
No significant circularity detected in derivation chain
full rationale
The abstract and context describe a new sliding-based propulsion mechanism in diatom chains, with claims of higher speed and efficiency at long wavelengths and specific aspect ratios. No equations, parameter fits, self-citations, or uniqueness theorems are provided that reduce any prediction or optimality result to the inputs by construction. The hydrodynamic model is presented as independent, with results derived from analysis or simulation rather than tautological redefinition or fitted-input renaming. This matches the default expectation for non-circular papers when no load-bearing reduction can be exhibited via quote and specific step.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Low Reynolds number hydrodynamics governs fluid interactions around the cell chains.
Reference graph
Works this paper leans on
-
[1]
Brennen and H
C. Brennen and H. Winet, Fluid mechanics of propulsion by cilia and flagella, Annual Review of Fluid Mechanics 9, 339 (1977)
1977
-
[2]
Gazzola, M
M. Gazzola, M. Argentina, and L. Mahadevan, Scaling macroscopic aquatic locomotion, Nature Physics10, 758 (2014)
2014
-
[3]
Childress,Mechanics of swimming and flying, 2 (Cam- bridge University Press, 1981)
S. Childress,Mechanics of swimming and flying, 2 (Cam- bridge University Press, 1981)
1981
-
[4]
E. M. Purcell, Life at low reynolds number, Physics and our world: reissue of the proceedings of a symposium in honor of Victor F Weisskopf , 47 (2014)
2014
-
[5]
Gray and G
J. Gray and G. J. Hancock, The propulsion of sea-urchin spermatozoa, Journal of Experimental Biology32, 802 (1955)
1955
-
[6]
Brokaw, Non-sinusoidal bending waves of sperm flag- ella, J
C. Brokaw, Non-sinusoidal bending waves of sperm flag- ella, J. exp. Biol43, 155 (1965)
1965
-
[7]
J. J. Higdon, A hydrodynamic analysis of flagellar propul- sion, Journal of Fluid Mechanics90, 685 (1979)
1979
-
[8]
Dresdner and D
R. Dresdner and D. Katz, Relationships of mammalian sperm motility and morphology to hydrodynamic aspects of cell function, Biology of reproduction25, 920 (1981)
1981
-
[9]
Tam and A
D. Tam and A. Hosoi, Optimal kinematics and morpholo- gies for spermatozoa, Physical Review E—Statistical, Nonlinear, and Soft Matter Physics83, 045303 (2011)
2011
-
[10]
Lauga and T
E. Lauga and T. R. Powers, The hydrodynamics of swim- ming microorganisms, Reports on progress in physics72, 096601 (2009)
2009
-
[11]
E. V. Armbrust, The life of diatoms in the world’s oceans, Nature459, 185 (2009)
2009
-
[12]
F. E. Round, R. M. Crawford, and D. G. Mann,Diatoms: biology and morphology of the genera(Cambridge univer- sity press, 1990). 9
1990
-
[13]
N. C. Poulsen, I. Spector, T. P. Spurck, T. F. Schultz, and R. Wetherbee, Diatom gliding is the result of an actin-myosin motility system, Cell motility and the cy- toskeleton44, 23 (1999)
1999
-
[14]
K. G. Bondoc-Naumovitz, E. Crosato, and K. Y. Wan, Functional morphology of gliding motility in benthic di- atoms, Proceedings of the National Academy of Sciences 122, e2426910122 (2025)
2025
-
[15]
Zhang, H
Q. Zhang, H. T. Leng, H. Li, K. R. Arrigo, and M. Prakash, Ice gliding diatoms establish record-low tem- perature limits for motility in a eukaryotic cell, Pro- ceedings of the National Academy of Sciences122, e2423725122 (2025)
2025
-
[16]
Yamaoka, Y
N. Yamaoka, Y. Suetomo, T. Yoshihisa, and S. Sonobe, Motion analysis and ultrastructural study of a colo- nial diatom, bacillaria paxillifer, Journal of Electron Mi- croscopy65, 211 (2016)
2016
-
[17]
O. F. M¨ uller,Kleine Schriften aus der Naturhistorie, vol. 1(Buchhandlung der Gelehrten, 1782)
-
[18]
A.-M. M. Schmid, The “paradox” diatom bacillaria pax- illifer (bacillariophyta) revisited 1, Journal of Phycology 43, 139 (2007)
2007
-
[19]
Harbich, Modeling the synchronization of the move- ment of bacillaria paxillifer by a kuramoto model with time delay, The Mathematical Biology of Diatoms , 193 (2023)
T. Harbich, Modeling the synchronization of the move- ment of bacillaria paxillifer by a kuramoto model with time delay, The Mathematical Biology of Diatoms , 193 (2023)
2023
-
[20]
S. E. Spagnolie and E. Lauga, The optimal elastic flagel- lum, Physics of Fluids22(2010)
2010
-
[21]
Lauga and C
E. Lauga and C. Eloy, Shape of optimal active flagella, Journal of Fluid Mechanics730, R1 (2013)
2013
-
[22]
Ishimoto, Hydrodynamic evolution of sperm swim- ming: Optimal flagella by a genetic algorithm, Journal of Theoretical Biology399, 166 (2016)
K. Ishimoto, Hydrodynamic evolution of sperm swim- ming: Optimal flagella by a genetic algorithm, Journal of Theoretical Biology399, 166 (2016)
2016
-
[23]
A. Das, M. Styslinger, D. M. Harris, and R. Zenit, Force and torque-free helical tail robot to study low reynolds number micro-organism swimming, Review of Scientific Instruments93(2022)
2022
-
[24]
W. Wang, Q. Liu, I. Tanasijevic, M. F. Reynolds, A. J. Cortese, M. Z. Miskin, M. C. Cao, D. A. Muller, A. C. Molnar, E. Lauga,et al., Cilia metasurfaces for electron- ically programmable microfluidic manipulation, Nature 605, 681 (2022)
2022
-
[25]
Brambilla, E
M. Brambilla, E. Ferrante, M. Birattari, and M. Dorigo, Swarm robotics: a review from the swarm engineering perspective, Swarm Intelligence7, 1 (2013)
2013
-
[26]
Gardi, S
G. Gardi, S. Ceron, W. Wang, K. Petersen, and M. Sitti, Microrobot collectives with reconfigurable morphologies, behaviors, and functions, Nature communications13, 2239 (2022)
2022
-
[27]
H. Xie, M. Sun, X. Fan, Z. Lin, W. Chen, L. Wang, L. Dong, and Q. He, Reconfigurable magnetic microrobot swarm: Multimode transformation, locomotion, and ma- nipulation, Science robotics4, eaav8006 (2019)
2019
-
[28]
X. Ju, C. Chen, C. M. Oral, S. Sevim, R. Golesta- nian, M. Sun, N. Bouzari, X. Lin, M. Urso, J. S. Nam, et al., Technology roadmap of micro/nanorobots, ACS nano (2025)
2025
-
[29]
iNaturalist contributor, Observation photo on inatural- ist,https://www.inaturalist.org/photos/11624974 (2017), accessed 2025-11-11
arXiv 2017
-
[30]
M. R. Kapinga and R. Gordon, Cell attachment in the motile colonial diatom bacillaria paxillifer, Diatom re- search7, 215 (1992)
1992
-
[31]
Balboa Usabiaga, B
F. Balboa Usabiaga, B. Kallemov, B. Delmotte, A. Bhalla, B. Griffith, and A. Donev, Hydrodynamics of suspensions of passive and active rigid particles: a rigid multiblob approach, Communications in Applied Math- ematics and Computational Science11, 217 (2017)
2017
-
[32]
Delmotte and F
B. Delmotte and F. B. Usabiaga, Modeling complex par- ticle suspensions: Perspectives on the rigid multiblob method, Physical Review Fluids10, 100701 (2025)
2025
-
[33]
F. B. Usabiaga and B. Delmotte, A numerical method for suspensions of articulated bodies in viscous flows, Journal of Computational Physics464, 111365 (2022)
2022
-
[34]
G. J. Hancock, The self-propulsion of microscopic organ- isms through liquids, Proceedings of the Royal Society of London. Series A. Mathematical and Physical Sciences 217, 96 (1953)
1953
-
[35]
R. G. Cox, The motion of long slender bodies in a viscous fluid part 1. general theory, Journal of Fluid mechanics 44, 791 (1970)
1970
-
[36]
Happel and H
J. Happel and H. Brenner,Low Reynolds number hydro- dynamics: with special applications to particulate media, Vol. 1 (Springer Science & Business Media, 2012)
2012
-
[37]
Fr´ ankov´ a-Koz´ akov´ a, P
M. Fr´ ankov´ a-Koz´ akov´ a, P. Marvan, and R. Geriˇ s, Halophilous diatoms in czech running waters: Pleurosira laevis and bacillaria paxillifera, inProceedings of the 1st Central European Diatom Meeting. Berlin, Germany (2007) pp. 39–44
2007
-
[38]
M. R. Kapinga and R. Gordon, Cell motility rhythms in bacillaria paxillifer, Diatom research7, 221 (1992)
1992
-
[39]
J. S. Guasto, R. Rusconi, and R. Stocker, Fluid mechan- ics of planktonic microorganisms, Annual Review of Fluid Mechanics44, 373 (2012)
2012
-
[40]
Chattopadhyay, R
S. Chattopadhyay, R. Moldovan, C. Yeung, and X. Wu, Swimming efficiency of bacterium escherichia coli, Pro- ceedings of the National Academy of Sciences103, 13712 (2006)
2006
-
[41]
Omori, H
T. Omori, H. Ito, and T. Ishikawa, Swimming microor- ganisms acquire optimal efficiency with multiple cilia, Proceedings of the National Academy of Sciences117, 30201 (2020)
2020
-
[42]
Schuech, T
R. Schuech, T. Hoehfurtner, D. J. Smith, and S. Humphries, Motile curved bacteria are pareto-optimal, Proceedings of the National Academy of Sciences116, 14440 (2019)
2019
-
[43]
Lovecchio, E
S. Lovecchio, E. Climent, R. Stocker, and W. M. Durham, Chain formation can enhance the vertical migration of phytoplankton through turbulence, Science advances5, eaaw7879 (2019)
2019
-
[44]
J. S. Font-Mu˜ noz, R. Jeanneret, J. Arrieta, S. Angl` es, A. Jordi, I. Tuval, and G. Basterretxea, Collective sink- ing promotes selective cell pairing in planktonic pennate diatoms, Proceedings of the National Academy of Sci- ences116, 15997 (2019)
2019
-
[45]
Moreau, Controllability and optimal control of mi- croswimmers: theory and applications, Journal of the Physical Society of Japan92, 121005 (2023)
C. Moreau, Controllability and optimal control of mi- croswimmers: theory and applications, Journal of the Physical Society of Japan92, 121005 (2023)
2023
-
[46]
Xiong, Z
T. Xiong, Z. Liu, Y. Wang, C. J. Ong, and L. Zhu, Chemotactic navigation in robotic swimmers via reset- free hierarchical reinforcement learning, Nature Commu- nications16, 5441 (2025)
2025
-
[47]
Y. Liu, Z. Wang, and A. C. Tsang, Reinforcement learn- ing selects multimodal locomotion strategies for bioin- spired microswimmers, Soft Matter21, 2363 (2025)
2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.