Physical Constraints on the Rhythmicity of the Biological Clock
Pith reviewed 2026-05-19 03:43 UTC · model grok-4.3
The pith
The KaiABC clock minimizes free energy cost at a natural ~21-hour period that entrains to 24-hour days when external forcing exceeds ~10% of metabolic rate.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
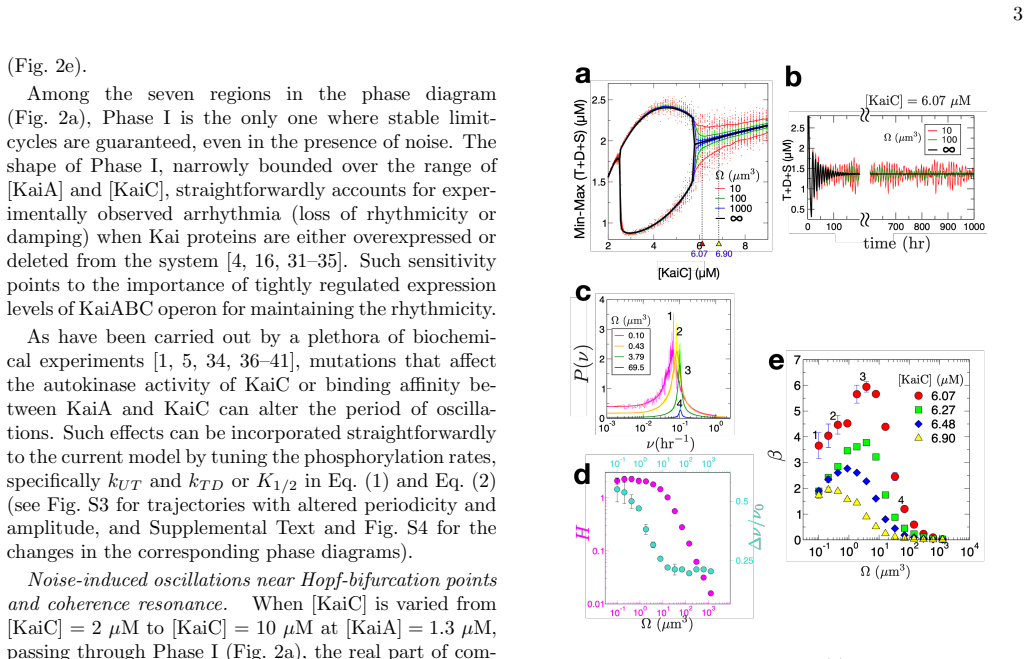
Within the oscillatory phase of the KaiABC system the cost-minimizing condition selected by the thermodynamic uncertainty relation produces an intrinsic ~21-hr rhythm. This rhythm entrains to 24-hr environmental signals as long as the forcing amplitude exceeds ~10 % of the metabolic rate. The phase diagram in KaiC and KaiA concentrations reveals a narrowly bounded oscillatory region, which accounts for loss of rhythmicity upon protein over-expression. An optimal level of intrinsic noise can induce oscillations even beyond the Hopf bifurcation, thereby expanding the oscillatory phase.
What carries the argument
The cost-minimizing condition obtained from the thermodynamic uncertainty relation applied to the stochastic KaiABC reaction network, which fixes the operating point that yields the ~21-hr intrinsic period.
If this is right
- The oscillatory phase occupies only a narrow interval in KaiC-KaiA concentration space, so overexpression of either protein disrupts rhythmicity.
- Higher free energy dissipation is required to achieve greater rhythmic precision because intrinsic noise is amplified in small reaction volumes.
- Entrainment to a 24-hr external cycle occurs once the forcing amplitude surpasses roughly 10 percent of the metabolic rate.
- An optimal amount of stochastic noise can generate oscillations even outside the deterministic Hopf bifurcation, widening the range of conditions that support rhythmicity.
Where Pith is reading between the lines
- Similar cost-precision trade-offs may set intrinsic periods in other circadian systems if the same thermodynamic relations govern their clocks.
- Cells could adjust total Kai protein levels to operate near the energy minimum while retaining robust entrainment to daily cycles.
- The framework predicts that altering noise levels through volume or reaction rates should shift the boundaries of the oscillatory phase in predictable ways.
Load-bearing premise
The thermodynamic uncertainty relation can be applied directly to the precision of the macroscopic rhythm produced by the stochastic KaiABC network, with total concentrations of KaiC and KaiA as the only relevant control parameters.
What would settle it
Direct measurement of free energy dissipated per cycle in reconstituted KaiABC oscillators tuned to different intrinsic periods, to test whether the minimum cost indeed lies near 21 hours.
Figures



read the original abstract
Circadian rhythms in living organisms are temporal orders emerging from biochemical circuits driven out of equilibrium. Here, considering the KaiABC system, a minimal model in the synthetic biology, we study how the oscillation emerges from the circuit made of three Kai proteins and ATP alone. The phase diagram constructed in terms of KaiC and KaiA concentrations reveals a narrowly bounded oscillatory phase, which naturally explains arrhythmia upon protein over-expression. As dictated by the cost-precision trade-offs of the thermodynamic uncertainty relations, the presence of intrinsic noise, amplified in small systems, demands higher free energy cost to achieve greater rhythmic precision. The cost-minimizing condition within the oscillatory phase is found to generate $\sim$21-hr rhythm, which is entrained to 24-hr environmental signals as long as the forcing amplitude is greater than $\sim 10$ \% of the metabolic rate. An optimal level of intrinsic noise can also induce oscillations even beyond the Hopf bifurcation, effectively expanding the oscillatory phase. Our study clarifies how the physical factors, such as regulatory mechanism, energy cost, and stochastic noise contribute to the operation of biological clocks.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents a minimal stochastic model of the KaiABC circadian clock consisting of three Kai proteins and ATP. It constructs a phase diagram in the plane of total KaiC and KaiA concentrations that identifies a narrow oscillatory regime explaining arrhythmia upon overexpression. Within this regime the authors invoke the thermodynamic uncertainty relation to argue that free-energy cost minimization under a precision constraint selects a ~21-hour period. This rhythm is reported to entrain to 24-hour environmental forcing provided the amplitude exceeds ~10% of the metabolic rate. Optimal intrinsic noise is also claimed to induce oscillations beyond the deterministic Hopf bifurcation.
Significance. If substantiated, the work would link stochastic-thermodynamic constraints to the period selection and robustness of a biological clock, offering a physical rationale for the narrow concentration window supporting rhythmicity and for the specific ~21-hour value that emerges under cost minimization. The numerical exploration of the stochastic model and the attempt to apply TUR to rhythm precision constitute the main technical contributions, though their impact is limited by the absence of direct experimental comparisons and by the need for a clearer derivation of the TUR adaptation.
major comments (2)
- [Cost-precision trade-off section] Cost-precision trade-off section: the claim that cost minimization within the oscillatory phase generates the ~21 hr rhythm rests on direct substitution of the period variance into the thermodynamic uncertainty relation. Standard TUR bounds apply to time-integrated currents or steady-state observables; the adaptation to the variance of return times (or phase diffusion constant) on a noisy limit cycle requires an explicit derivation via the Fokker-Planck description of the phase variable or large-deviation theory. The manuscript does not provide this step or cite a specific result justifying the substitution, which is load-bearing for the central period-selection result.
- [Entrainment analysis] Entrainment analysis: the threshold forcing amplitude of ~10% of the metabolic rate for entrainment to 24 hr signals is stated without a precise definition of the metabolic rate in the model equations and without reported variability or error bars from the stochastic trajectories. This renders the quantitative threshold difficult to assess and weakens the entrainment claim.
minor comments (3)
- [Abstract] The abstract describes the model as belonging to 'synthetic biology' while it is a minimal representation of the natural KaiABC system; this phrasing should be clarified.
- [Simulation details] No error bars or standard deviations are shown for periods or thresholds extracted from stochastic simulations, and a table of all rate constants and total concentrations used is absent, limiting reproducibility.
- [Phase diagram figure] The phase diagram would be clearer if the deterministic Hopf bifurcation boundary were overlaid on the stochastic oscillatory region.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive comments, which have helped us improve the manuscript. We address each major comment below and have revised the manuscript to strengthen the derivations and clarify the quantitative claims.
read point-by-point responses
-
Referee: [Cost-precision trade-off section] Cost-precision trade-off section: the claim that cost minimization within the oscillatory phase generates the ~21 hr rhythm rests on direct substitution of the period variance into the thermodynamic uncertainty relation. Standard TUR bounds apply to time-integrated currents or steady-state observables; the adaptation to the variance of return times (or phase diffusion constant) on a noisy limit cycle requires an explicit derivation via the Fokker-Planck description of the phase variable or large-deviation theory. The manuscript does not provide this step or cite a specific result justifying the substitution, which is load-bearing for the central period-selection result.
Authors: We agree that a direct substitution of the period variance into the standard TUR requires explicit justification for a noisy limit cycle. In the revised manuscript we have added a dedicated subsection deriving the relevant bound from the Fokker-Planck equation for the phase variable, showing that the phase diffusion constant satisfies a TUR-like inequality whose equality case yields the cost-minimizing period. We also cite the appropriate stochastic-thermodynamics literature on oscillators. This addition makes the central period-selection argument self-contained. revision: yes
-
Referee: [Entrainment analysis] Entrainment analysis: the threshold forcing amplitude of ~10% of the metabolic rate for entrainment to 24 hr signals is stated without a precise definition of the metabolic rate in the model equations and without reported variability or error bars from the stochastic trajectories. This renders the quantitative threshold difficult to assess and weakens the entrainment claim.
Authors: We accept that the entrainment threshold must be defined more precisely. In the revised version we explicitly identify the metabolic rate with the time-averaged ATP hydrolysis flux in the unforced oscillatory state (computed directly from the model rate constants). We have also added error bars and variability statistics obtained from ensembles of stochastic trajectories, confirming that the entrainment transition remains near 10% with quantified uncertainty. revision: yes
Circularity Check
No significant circularity; derivation remains self-contained
full rationale
The manuscript builds a phase diagram in KaiC/KaiA concentration space from the underlying reaction network, identifies an oscillatory regime, and then invokes the thermodynamic uncertainty relation to relate free-energy dissipation rate to rhythmic precision. The cost-minimizing condition inside that regime is reported to select a ~21 h period, after which entrainment to a 24 h drive is examined as a function of forcing amplitude. These steps rest on explicit dynamical equations and the standard TUR inequality applied to a time-integrated current; no equation is shown to be identical to its own input by construction, no parameter is fitted on a subset and then relabeled a prediction, and no load-bearing uniqueness claim is imported solely via self-citation. The derivation therefore retains independent content from the physical model and the cost-precision trade-off.
Axiom & Free-Parameter Ledger
free parameters (2)
- KaiC and KaiA total concentrations
- Forcing amplitude threshold (~10 % of metabolic rate)
axioms (1)
- domain assumption Thermodynamic uncertainty relations apply directly to the precision of the emergent macroscopic rhythm
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The TUR for periodic dynamics can be written as Q = ΔS_tot / k_B * <δT_os²> / <T_os>² = Ṡ_tot / k_B * <δT_os²> / <T_os>² ≥ 2
-
IndisputableMonolith/Foundation/AlphaCoordinateFixation.leanJ_uniquely_calibrated_via_higher_derivative unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
cost-minimizing condition within the oscillatory phase is found to generate ∼21-hr rhythm
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
M. Nakajima, K. Imai, H. Ito, T. Nishiwaki, Y. Mu- rayama, H. Iwasaki, T. Oyama, and T. Kondo, Reconsti- tution of Circadian Oscillation of Cyanobacterial KaiC Phosphorylation in Vitro, Science 308, 414 (2005)
work page 2005
-
[2]
H. Kageyama, T. Nishiwaki, M. Nakajima, H. Iwasaki, T. Oyama, and T. Kondo, Cyanobacterial circadian pace- maker: Kai protein complex dynamics in the KaiC phos- phorylation cycle in vitro, Molecular Cell 23, 161 (2006)
work page 2006
-
[3]
K. Terauchi, Y. Kitayama, T. Nishiwaki, K. Miwa, Y. Murayama, T. Oyama, and T. Kondo, ATPase ac- tivity of KaiC determines the basic timing for circadian clock of cyanobacteria, Proc. Natl. Acad. Sci. 104, 16377 (2007)
work page 2007
-
[4]
M. Ishiura, S. Kutsuna, S. Aoki, H. Iwasaki, C. R. An- dersson, A. Tanabe, S. S. Golden, C. H. Johnson, and T. Kondo, Expression of a Gene Cluster kaiABC as a Cir- cadian Feedback Process in Cyanobacteria, Science 281, 1519 (1998)
work page 1998
- [5]
-
[6]
V. Dvornyk, O. Vinogradova, and E. Nevo, Origin and evolution of circadian clock genes in prokaryotes, Proc. Natl. Acad. Sci. 100, 2495 (2003)
work page 2003
-
[7]
Y. Xu, T. Mori, R. Pattanayek, S. Pattanayek, M. Egli, and C. H. Johnson, Identification of key phosphorylation sites in the circadian clock protein KaiC by crystallo- graphic and mutagenetic analyses, Proc. Natl. Acad. Sci. 101, 13933 (2004)
work page 2004
- [8]
-
[9]
J. A. Swan, C. R. Sandate, A. G. Chavan, A. M. Free- berg, D. Etwaru, D. C. Ernst, J. G. Palacios, S. S. Golden, A. LiWang, G. C. Lander, and C. L. Partch, Cou- pling of distant ATPase domains in the circadian clock protein KaiC, Nat. Struct. Mol. Biol. 29, 759 (2022)
work page 2022
-
[10]
Y. Furuike, Y. Onoue, S. Saito, T. Mori, and S. Akiyama, The priming phosphorylation of KaiC is activated by the release of its autokinase autoinhibition, PNAS nexus 4, pgaf136 (2025)
work page 2025
-
[11]
M. Sasai, Mechanism of autonomous synchronization of the circadian KaiABC rhythm, Scientific Reports 11, 4713 (2021)
work page 2021
-
[12]
M. Sasai, Role of the reaction-structure coupling in tem- perature compensation of the KaiABC circadian rhythm, PLOS Computational Biology 18, e1010494 (2022)
work page 2022
-
[13]
J. S. van Zon, D. K. Lubensky, P. R. Altena, and P. R. ten Wolde, An allosteric model of circadian KaiC phos- phorylation, Proc. Natl. Acad. Sci. 104, 7420 (2007)
work page 2007
-
[14]
Sparrow, The Lorenz equations , Vol
C. Sparrow, The Lorenz equations , Vol. 109 (Springer New York, 1982)
work page 1982
-
[15]
Ott, Chaos in dynamical systems (Cambridge univer- sity press, 2002)
E. Ott, Chaos in dynamical systems (Cambridge univer- sity press, 2002)
work page 2002
-
[16]
M. J. Rust, J. S. Markson, W. S. Lane, D. S. Fisher, and E. K. O’Shea, Ordered phosphorylation governs oscilla- tion of a three-protein circadian clock, Science 318, 809 (2007)
work page 2007
-
[17]
E. Mara˜ n´ on, C. Fern´ andez-Gonz´ alez, and G. A. Tarran, Effect of temperature, nutrients and growth rate on pi- cophytoplankton cell size across the atlantic ocean, Sci- entific Reports 14, 28034 (2024)
work page 2024
-
[18]
E. J. Carpenter, A. Subramaniam, and D. G. Capone, Biomass and primary productivity of the cyanobacterium Trichodesmium spp. in the tropical N Atlantic ocean, Deep Sea Research Part I: Oceanographic Research Pa- pers 51, 173 (2004)
work page 2004
-
[19]
A. Hahn, M. Stevanovic, O. Mirus, and E. Schleiff, The TolC-like Protein HgdD of the Cyanobacterium An- abaena sp. PCC 7120 Is Involved in Secondary Metabo- lite Export and Antibiotic Resistance, J. Biol. Chem. 287, 41126 (2012)
work page 2012
-
[20]
H. Gang, T. Ditzinger, C.-Z. Ning, and H. Haken, Stochastic resonance without external periodic force, Phys. Rev. Lett. 71, 807 (1993)
work page 1993
-
[21]
A. S. Pikovsky and J. Kurths, Coherence resonance in a noise-driven excitable system, Phys. Rev. Lett. 78, 775 (1997)
work page 1997
-
[22]
O. Ushakov, H.-J. W¨ unsche, F. Henneberger, I. Kho- vanov, L. Schimansky-Geier, and M. Zaks, Coherence resonance near a Hopf bifurcation, Phys. Rev. Lett. 95, 123903 (2005)
work page 2005
-
[23]
Y. Cao, H. Wang, Q. Ouyang, and Y. Tu, The free-energy cost of accurate biochemical oscillations, Nat. Phys. 11, 772 (2015)
work page 2015
-
[24]
A. C. Barato and U. Seifert, Thermodynamic Uncer- tainty Relation for Biomolecular Processes, Phys. Rev. Lett. 114, 158101 (2015)
work page 2015
-
[25]
R. Marsland III, W. Cui, and J. M. Horowitz, The ther- modynamic uncertainty relation in biochemical oscilla- tions, J. R. Soc. Interface 16, 20190098 (2019)
work page 2019
-
[26]
M. Egli, R. Pattanayek, J. H. Sheehan, Y. Xu, T. Mori, J. A. Smith, and C. H. Johnson, Loop–loop interac- tions regulate KaiA-stimulated KaiC phosphorylation in the cyanobacterial KaiABC circadian clock, Biochem- istry 52, 1208 (2013)
work page 2013
-
[27]
F. Hayashi, H. Ito, M. Fujita, R. Iwase, T. Uzu- maki, and M. Ishiura, Stoichiometric interactions be- tween cyanobacterial clock proteins KaiA and KaiC, Biochem. Biophys. Res. Commun. 316, 195 (2004)
work page 2004
-
[28]
C. Brettschneider, R. J. Rose, S. Hertel, I. M. Axmann, A. J. Heck, and M. Kollmann, A sequestration feed- back determines dynamics and temperature entrainment of the KaiABC circadian clock, Mol. Syst. Biol. 6, 389 6 (2010)
work page 2010
-
[29]
D. T. Gillespie, Exact stochastic simulation of coupled chemical reactions, J. Phys. Chem. 81, 2340 (1977)
work page 1977
-
[30]
N. G. van Kampen, Stochastic Processes in Chem- istry and Physics (Elsevier, North Holland, Amsterdam, 2007)
work page 2007
-
[31]
S. E. Cohen, M. L. Erb, J. Selimkhanov, G. Dong, J. Hasty, J. Pogliano, and S. S. Golden, Dynamic lo- calization of the cyanobacterial circadian clock proteins, Current Biology 24, 1836 (2014)
work page 2014
-
[32]
N. Kawamoto, H. Ito, I. T. Tokuda, and H. Iwasaki, Damped circadian oscillation in the absence of KaiA in Synechococcus, Nature Commun. 11, 2242 (2020)
work page 2020
-
[33]
Y. Xu, T. Mori, and C. H. Johnson, Circadian clock- protein expression in cyanobacteria: rhythms and phase setting, EMBO J. (2000)
work page 2000
-
[34]
H. Iwasaki, T. Nishiwaki, Y. Kitayama, M. Nakajima, and T. Kondo, KaiA-stimulated KaiC phosphorylation in circadian timing loops in cyanobacteria, Proc. Natl. Acad. Sci. 99, 15788 (2002)
work page 2002
-
[35]
X. Qin, M. Byrne, T. Mori, P. Zou, D. R. Williams, H. Mchaourab, and C. H. Johnson, Intermolecular associ- ations determine the dynamics of the circadian KaiABC oscillator, Proc. Natl. Acad. Sci. 107, 14805 (2010)
work page 2010
-
[36]
Y. Kitayama, H. Iwasaki, T. Nishiwaki, and T. Kondo, KaiB functions as an attenuator of KaiC phosphorylation in the cyanobacterial circadian clock system, EMBO J. 22, 2127 (2003)
work page 2003
-
[37]
Y.-G. Chang, N.-W. Kuo, R. Tseng, and A. LiWang, Flexibility of the C-terminal, or CII, ring of KaiC governs the rhythm of the circadian clock of cyanobacteria, Proc. Natl. Acad. Sci. 108, 14431 (2011)
work page 2011
-
[38]
Y. Yunoki, K. Ishii, M. Yagi-Utsumi, R. Murakami, S. Uchiyama, H. Yagi, and K. Kato, ATP hydrolysis by KaiC promotes its KaiA binding in the cyanobac- terial circadian clock system, Life Science Alliance 2, 10.26508/lsa.201900368 (2019)
-
[39]
H. Nishimura, Y. Nakahira, K. Imai, A. Tsuruhara, H. Kondo, H. Hayashi, M. Hirai, H. Saito, and T. Kondo, Mutations in KaiA, a clock protein, extend the period of circadian rhythm in the cyanobacterium Synechococcus elongatus PCC 7942, Microbiology 148, 2903 (2002)
work page 2002
-
[40]
K. Ito-Miwa, Y. Furuike, S. Akiyama, and T. Kondo, Tuning the circadian period of cyanobacteria up to 6.6 days by the single amino acid substitutions in KaiC, Proc. Natl. Acad. Sci. 117, 20926 (2020)
work page 2020
-
[41]
R. Pattanayek, D. R. Williams, S. Pattanayek, Y. Xu, T. Mori, C. H. Johnson, P. L. Stewart, and M. Egli, Analysis of KaiA–KaiC protein interactions in the cyano- bacterial circadian clock using hybrid structural meth- ods, EMBO J 25, 2017 (2006)
work page 2017
-
[42]
T. R. Gingrich and J. M. Horowitz, Fundamental bounds on first passage time fluctuations for currents, Phys. Rev. Lett. 119, 170601 (2017)
work page 2017
- [43]
-
[44]
D. A. Beard and H. Qian, Chemical Biophysics: Quanti- tative Analysis of Cellular Systems (Cambridge Univer- sity Press, 2008)
work page 2008
-
[45]
M. Fang, A. G. Chavan, A. LiWang, and S. S. Golden, Synchronization of the circadian clock to the environ- ment tracked in real time, Proc. Natl. Acad. Sci. 120, e2221453120 (2023)
work page 2023
- [46]
-
[47]
A. Pikovsky, M. Rosenblum, and J. Kurths, Synchroniza- tion: a universal concept in nonlinear science , edited by B. Chirikov, C. Predrag, F. Moss, and H. Swinney (Cam- bridge University Press, 2002)
work page 2002
-
[48]
M. L. Heltberg, S. Krishna, L. P. Kadanoff, and M. H. Jensen, A tale of two rhythms: Locked clocks and chaos in biology, Cell Systems 12, 291 (2021)
work page 2021
-
[49]
W. Hwang and C. Hyeon, Energetic costs, precision, and transport efficiency of molecular motors, J. Phys. Chem. Lett. 9, 513 (2018)
work page 2018
-
[50]
Y. Song and C. Hyeon, Thermodynamic uncertainty re- lation to assess biological processes, J. Chem. Phys. 154, 130901 (2021)
work page 2021
- [51]
-
[52]
A. Murugan, D. A. Huse, and S. Leibler, Speed, dissipa- tion, and error in kinetic proofreading, Proc. Natl. Acad. Sci. 109, 12034 (2012)
work page 2012
-
[53]
Y. Song and C. Hyeon, Thermodynamic cost, speed, fluc- tuations, and error reduction of biological copy machines, J. Phys. Chem. Lett. 11, 3136 (2020)
work page 2020
-
[54]
J. D. Mallory, O. A. Igoshin, and A. B. Kolomeisky, Do we understand the mechanisms used by biological sys- tems to correct their errors?, J. Phys. Chem. B124, 9289 (2020)
work page 2020
-
[55]
Y. Song and C. Hyeon, Cost-precision trade-off relation determines the optimal morphogen gradient for accurate biological pattern formation, Elife 10, e70034 (2021)
work page 2021
- [56]
-
[57]
A. T. Winfree, The geometry of biological time , Vol. 12 (Springer Science & Business Media, 2001)
work page 2001
-
[58]
Kuramoto, Chemical oscillations, waves, and turbu- lence (Courier Corporation, 2003)
Y. Kuramoto, Chemical oscillations, waves, and turbu- lence (Courier Corporation, 2003)
work page 2003
-
[59]
Y. Song, J. O. Park, L. Tanner, Y. Nagano, J. D. Rabinowitz, and S. Y. Shvartsman, Energy budget of Drosophila embryogenesis, Current Biology 29, R566 (2019)
work page 2019
-
[60]
X. Yang, M. Heinemann, J. Howard, G. Huber, S. Iyer- Biswas, G. Le Treut, M. Lynch, K. L. Montooth, D. J. Needleman, S. Pigolotti, et al. , Physical bioenergetics: Energy fluxes, budgets, and constraints in cells, Proc. Natl. Acad. Sci. 118, e2026786118 (2021)
work page 2021
-
[61]
I. Di Terlizzi, M. Gironella, D. Herraez-Aguilar, T. Betz, F. Monroy, M. Baiesi, and F. Ritort, Variance sum rule for entropy production, Science 383, 971 (2024)
work page 2024
-
[62]
Z. Padamsey and N. L. Rochefort, Paying the brain’s energy bill, Curr. Opin. Neurobiol. 78, 102668 (2023)
work page 2023
-
[63]
S. D. Jamadar, A. Behler, H. Deery, and M. Breakspear, The metabolic costs of cognition, Trends in Cognitive Sci. (2025)
work page 2025
-
[64]
M. B. Elowitz and S. Leibler, A synthetic oscillatory network of transcriptional regulators, Nature 403, 335 (2000)
work page 2000
-
[65]
J. Stricker, S. Cookson, M. R. Bennett, W. H. Mather, L. S. Tsimring, and J. Hasty, A fast, robust and tunable synthetic gene oscillator, Nature 456, 516 (2008)
work page 2008
-
[66]
L. Potvin-Trottier, N. D. Lord, G. Vinnicombe, and J. Paulsson, Synchronous long-term oscillations in a syn- thetic gene circuit, Nature 538, 514 (2016). 7
work page 2016
-
[67]
P. Dong, Y. Fan, J. Sun, M. Lv, M. Yi, X. Tan, and S. Liu, A dynamic interaction process between KaiA and KaiC is critical to the cyanobacterial circadian oscillator, Scientific Reports 6, 25129 (2016). 8 SUPPLEMENTAL MATERIALS Linear stability analysis Dynamical systems ˙x = ⃗F (x) in 3D, expanded around a fixed point x∗ = (x∗, y∗, z∗) satisfying Fi(x∗, ...
work page 2016
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.