Modelling chronic stress as an excitatory-inhibitory perturbation in recurrent working-memory networks
Pith reviewed 2026-06-29 00:47 UTC · model grok-4.3
The pith
Stronger inhibitory-to-excitatory synapses recover all three signatures of chronic stress in working-memory networks.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
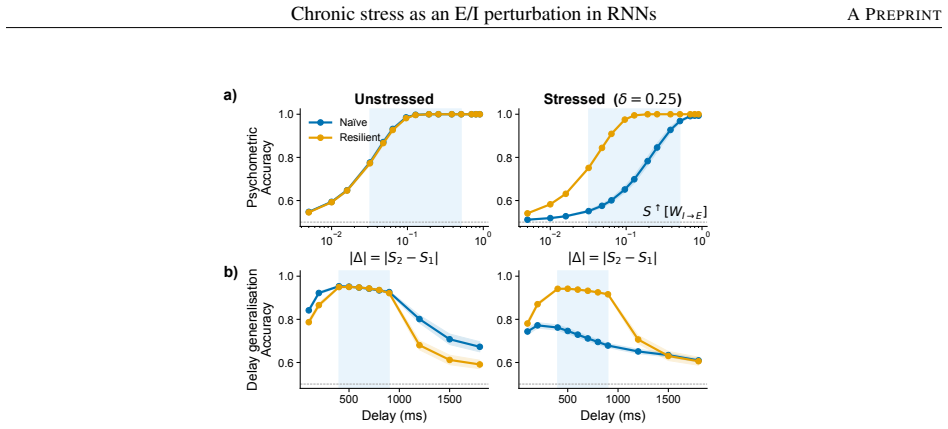
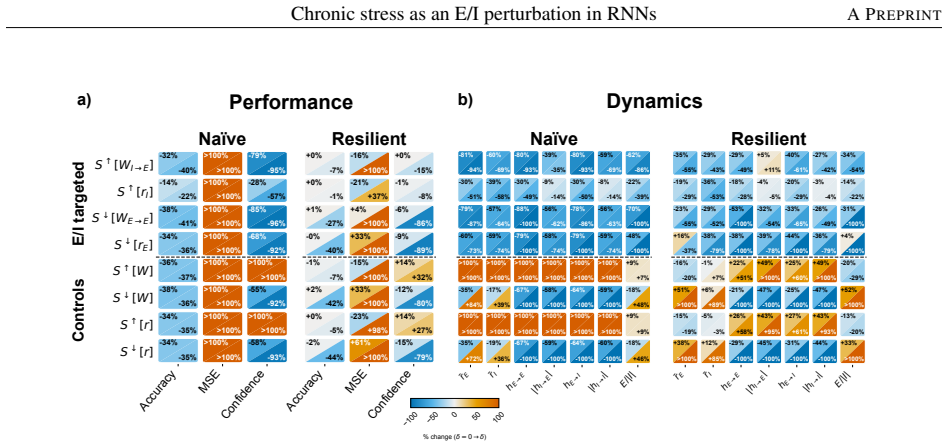
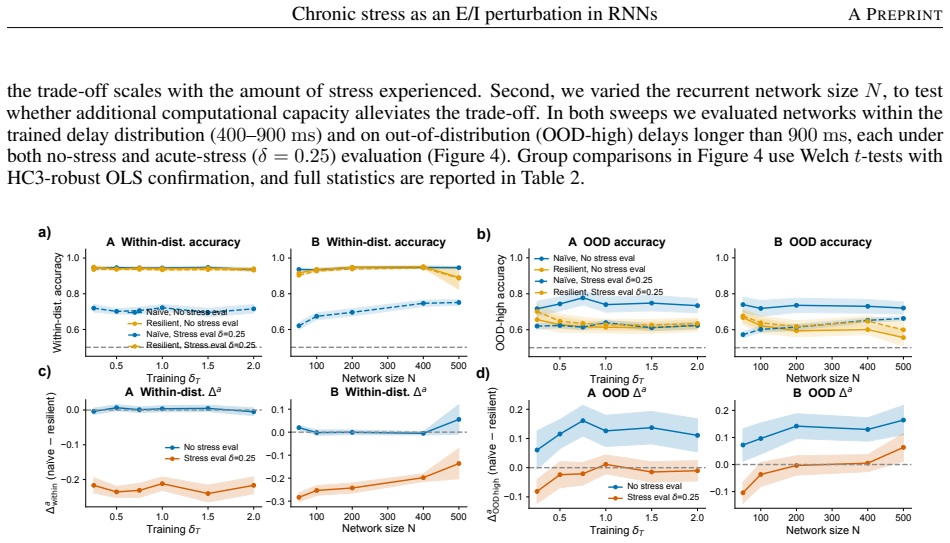
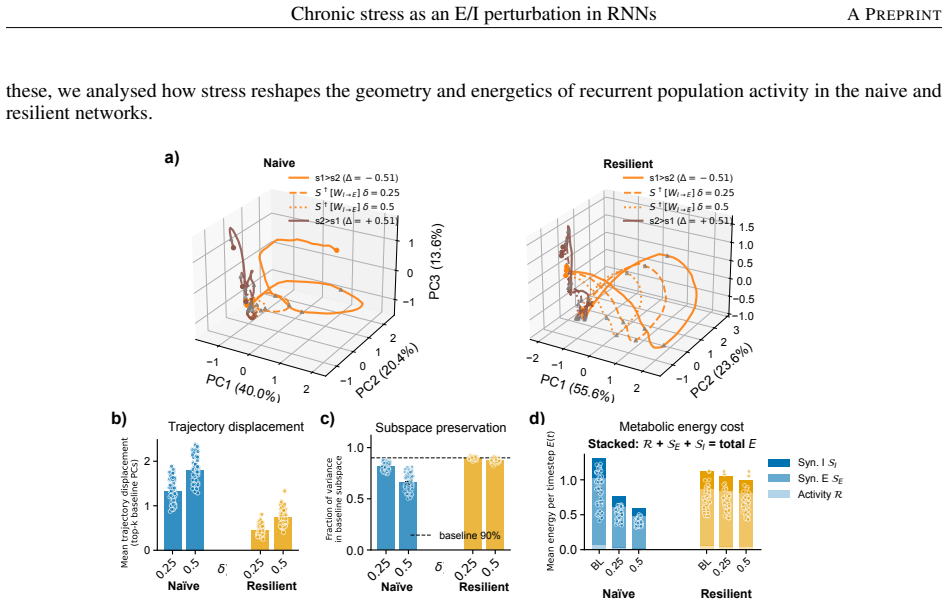
Among eight candidate synaptic or activity modulations, only stronger inhibitory-to-excitatory synapses simultaneously produce inhibitory dominance, excitatory hypofunction, and impaired working-memory performance. Networks trained under this mechanism maintain performance and remain in the same dynamical subspace and energetic regime with or without stress, yet show reduced generalization when the task demands longer memory intervals than those seen during training; the resilience-generalization trade-off holds across stress levels and network sizes.
What carries the argument
Recurrent networks trained on a working-memory task, with eight candidate operators that modulate synaptic strength or neuronal activity to model chronic stress.
If this is right
- Resilient networks preserve task performance under stress and stay within the same dynamical subspace and energetic regime.
- Resilient networks generalize less well to working-memory tasks that require longer retention intervals than those used in training.
- The resilience-generalization trade-off remains across different stress magnitudes and network sizes.
- Resilience training produces a more specialized solution tuned to the trained regime.
Where Pith is reading between the lines
- The model predicts that chronic stress biases circuits toward rigid, less adaptable solutions that resemble habit-like behavior.
- Interventions that selectively weaken inhibitory-to-excitatory synapses could restore both performance and flexibility after stress exposure.
- Similar stress operators could be tested in other recurrent circuits to see whether the same mechanism explains dysfunction outside working memory.
- The observed trade-off suggests a computational reason why stressed animals show reduced behavioral flexibility on novel problems.
Load-bearing premise
The three experimental signatures plus the working-memory task are enough to single out the right mechanism among the eight candidates.
What would settle it
Finding that biological prefrontal circuits under chronic stress do not show strengthened inhibitory-to-excitatory synapses while still displaying the three signatures, or that another of the eight operators matches the signatures equally well under additional biological constraints.
Figures

read the original abstract
Stress is an adaptive response coordinated by neural and physiological systems. While acute stress can enhance survival, chronic stress drives structural brain changes, cognitive dysfunction, and increased psychiatric risk. At the cellular level, chronic stress shifts the excitatory-inhibitory (E/I) balance of prefrontal pyramidal neurons toward inhibitory dominance, yet the mechanisms underlying these alterations are still unknown. We here investigate possible mechanisms causing inhibitory dominance using recurrent neuronal networks trained on a working memory task. Chronic stress is modelled as a modulation in synaptic strength or neuronal activity, systematically comparing eight candidate operators against three experimentally motivated signatures of stress-induced prefrontal dysfunction: inhibitory dominance, excitatory hypofunction, and impaired task performance. These signatures are all recovered by a single stress mechanism, stronger inhibitory-to-excitatory synapses. Contrasting naive networks with resilient networks trained under the stress mechanism, we find that resilience training not only preserves task performance under stress, but also confines the network to the same dynamical subspace and energetic regime with and without stress. This resilience comes at a cost: resilient networks generalise less well when the task requires longer memory than seen during training, indicating that resilient networks find a specialised solution tuned to the trained regime. This trade-off between resilience and generalization performance persists across stress magnitude and network size, offering a computational analogue of the shift toward rigid, habit-like behaviour reported in animal following chronic stress.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper models chronic stress as one of eight candidate E/I perturbations (synaptic strength or activity modulations) in recurrent networks trained on a working-memory task. It reports that only stronger inhibitory-to-excitatory synapses simultaneously reproduce the three experimental signatures of inhibitory dominance, excitatory hypofunction, and impaired task performance. Networks trained under this mechanism are then shown to be resilient (preserving performance, dynamical subspace, and energetic regime under stress) while exhibiting reduced generalization to longer memory delays, with the resilience-generalization trade-off persisting across stress magnitudes and network sizes.
Significance. If the central identification holds, the work supplies a mechanistic account linking a specific synaptic operator to multiple stress-induced prefrontal phenotypes and supplies a computational analogue for the resilience-rigidity trade-off observed in stressed animals. The systematic enumeration of eight operators and the subsequent analysis of subspace confinement and generalization cost are positive features that go beyond single-mechanism fitting.
major comments (1)
- [Results, operator comparison] Results (mechanism comparison): the uniqueness claim—that only the I-to-E strengthening operator recovers all three signatures while the other seven do not—rests on the assumption that the three signatures supply independent constraints. Because inhibitory dominance and excitatory hypofunction are both direct, correlated consequences of any E/I shift in the same recurrent circuit, and task impairment follows from the resulting dynamics, the reported analysis does not demonstrate that the signatures are sufficiently orthogonal to exclude alternative operators that could be tuned to match the same three observables. A quantitative measure of signature independence or an additional, biologically motivated signature would be required to support the uniqueness conclusion.
minor comments (2)
- [Abstract / Methods] The abstract and methods should explicitly state the precise definitions of the eight operators (e.g., which synapses are scaled and by what functional form) so that the comparison can be reproduced without ambiguity.
- [Figures] Figure legends for the resilience and generalization panels should report the exact number of networks, random seeds, and statistical tests used to support the claim that the trade-off 'persists across stress magnitude and network size.'
Simulated Author's Rebuttal
We thank the referee for their constructive comments, which help clarify the strength of our uniqueness claim. We address the major comment below.
read point-by-point responses
-
Referee: Results (mechanism comparison): the uniqueness claim—that only the I-to-E strengthening operator recovers all three signatures while the other seven do not—rests on the assumption that the three signatures supply independent constraints. Because inhibitory dominance and excitatory hypofunction are both direct, correlated consequences of any E/I shift in the same recurrent circuit, and task impairment follows from the resulting dynamics, the reported analysis does not demonstrate that the signatures are sufficiently orthogonal to exclude alternative operators that could be tuned to match the same three observables. A quantitative measure of signature independence or an additional, biologically motivated signature would be required to support the uniqueness conclusion.
Authors: We agree that inhibitory dominance and excitatory hypofunction are mechanistically linked through E/I balance and that task impairment is a downstream consequence. However, the eight operators represent distinct biological perturbations (specific synaptic weight changes vs. activity modulations), and our systematic parameter sweeps show that only I-to-E strengthening simultaneously matches the quantitative experimental signatures (direction and magnitude of E/I shift plus performance drop) reported in the stress literature. Other operators either produce mismatched E/I ratios, fail to impair performance at observed levels, or require implausible parameter values outside biological ranges. While the signatures are not fully orthogonal, their combination still discriminates among the operators in our enumeration. We will add a supplementary figure quantifying pairwise correlations among the three signatures across all operators and a discussion paragraph addressing the referee's concern about independence. This constitutes a partial revision. revision: partial
Circularity Check
No significant circularity; mechanism identification uses independent experimental signatures
full rationale
The paper compares eight candidate operators against three experimentally motivated signatures (inhibitory dominance, excitatory hypofunction, impaired task performance) drawn from external literature. The identification of stronger inhibitory-to-excitatory synapses as the sole matching mechanism does not reduce to a self-definitional loop, a fitted parameter renamed as prediction, or a self-citation chain. Resilience and generalization results are obtained by direct simulation of networks trained under the identified mechanism; they are consequences rather than inputs. No equations or steps in the provided derivation exhibit the enumerated circularity patterns. The analysis remains self-contained against the external signatures.
Axiom & Free-Parameter Ledger
free parameters (1)
- perturbation strength
axioms (1)
- domain assumption Recurrent networks trained on a working memory task adequately capture prefrontal E/I balance relevant to stress-induced dysfunction.
Reference graph
Works this paper leans on
-
[1]
A comprehensive overview on stress neurobiology: Basic concepts and clinical implications
Lívea Dornela Godoy, Matheus Teixeira Rossignoli, Polianna Delfino-Pereira, Norberto Garcia-Cairasco, and Eduardo Henrique de Lima Umeoka. A comprehensive overview on stress neurobiology: Basic concepts and clinical implications. 12:127. doi:10.3389/fnbeh.2018.00127. URL http://dx.doi.org/10.3389/fnbeh. 2018.00127
-
[2]
Katharine Ann James, Juliet Ilena Stromin, Nina Steenkamp, and Marc Irwin Combrinck. Understand- ing the relationships between physiological and psychosocial stress, cortisol and cognition. 14:1085950. doi:10.3389/fendo.2023.1085950. URLhttp://dx.doi.org/10.3389/fendo.2023.1085950. 18 Chronic stress as an E/I perturbation in RNNsA PREPRINT
-
[3]
Stress signalling pathways that impair prefrontal cortex structure and function
Amy F T Arnsten. Stress signalling pathways that impair prefrontal cortex structure and function. 10:410–422. doi:10.1038/nrn2648. URLhttp://dx.doi.org/10.1038/nrn2648
-
[4]
Neurocognitive effects of stress: a metaparadigm perspective
Eun Joo Kim and Jeansok J Kim. Neurocognitive effects of stress: a metaparadigm perspective. 28:2750–2763. doi:10.1038/s41380-023-01986-4. URLhttp://dx.doi.org/10.1038/s41380-023-01986-4
-
[5]
David M Devilbiss, Rick L Jenison, and Craig W Berridge. Stress-induced impairment of a working memory task: role of spiking rate and spiking history predicted discharge. 8:e1002681. doi:10.1371/journal.pcbi.1002681. URL http://dx.doi.org/10.1371/journal.pcbi.1002681
-
[6]
Chronic stress causes frontostriatal reorganization and affects decision-making
Eduardo Dias-Ferreira, João C Sousa, Irene Melo, Pedro Morgado, Ana R Mesquita, João J Cerqueira, Rui M Costa, and Nuno Sousa. Chronic stress causes frontostriatal reorganization and affects decision-making. 325: 621–625. doi:10.1126/science.1171203. URLhttp://dx.doi.org/10.1126/science.1171203
-
[7]
Effects of chronic stress on cognitive function - from neurobiology to intervention
Milena Girotti, Sarah E Bulin, and Flavia R Carreno. Effects of chronic stress on cognitive function - from neurobiology to intervention. 33:100670. doi:10.1016/j.ynstr.2024.100670. URL http://dx.doi.org/10. 1016/j.ynstr.2024.100670
-
[8]
Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex
Bruce S McEwen, Carla Nasca, and Jason D Gray. Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. 41:3–23. doi:10.1038/npp.2015.171. URL http://dx.doi.org/10.1038/npp.2015. 171
-
[9]
Margaret T Davis, Sophie E Holmes, Robert H Pietrzak, and Irina Esterlis. Neurobiology of chronic stress-related psychiatric disorders: Evidence from molecular imaging studies. 1:247054701771091. doi:10.1177/2470547017710916. URLhttp://dx.doi.org/10.1177/2470547017710916
-
[10]
The evolving neurobiology of early-life stress
Matthew T Birnie and Tallie Z Baram. The evolving neurobiology of early-life stress. 113:1474–1490. doi:10.1016/j.neuron.2025.02.016. URLhttp://dx.doi.org/10.1016/j.neuron.2025.02.016
-
[11]
Influence of glutamate and GABA transport on brain excita- tory/inhibitory balance
Sheila Ms Sears and Sandra J Hewett. Influence of glutamate and GABA transport on brain excita- tory/inhibitory balance. 246:1069–1083. doi:10.1177/1535370221989263. URL http://dx.doi.org/10. 1177/1535370221989263
-
[12]
Keeping excitation-inhibition ratio in balance
Sergei Kirischuk. Keeping excitation-inhibition ratio in balance. 23:5746. doi:10.3390/ijms23105746. URL http://dx.doi.org/10.3390/ijms23105746
-
[13]
Yu-Ting Hu, Zhong-Lin Tan, Dusan Hirjak, and Georg Northoff. Brain-wide changes in excitation-inhibition balance of major depressive disorder: a systematic review of topographic patterns of GABA- and glutamater- gic alterations. 28:3257–3266. doi:10.1038/s41380-023-02193-x. URL http://dx.doi.org/10.1038/ s41380-023-02193-x
-
[14]
Chloe E Page and Laurence Coutellier. Prefrontal excitatory/inhibitory balance in stress and emotional disorders: Evidence for over-inhibition. 105:39–51. doi:10.1016/j.neubiorev.2019.07.024. URL http://dx.doi.org/10. 1016/j.neubiorev.2019.07.024
-
[15]
Jessica M McKlveen, Rachel L Morano, Maureen Fitzgerald, Sandra Zoubovsky, Sarah N Cassella, Jessie R Scheimann, Sriparna Ghosal, Parinaz Mahbod, Benjamin A Packard, Brent Myers, Mark L Baccei, and James P Herman. Chronic stress increases prefrontal inhibition: A mechanism for stress-induced prefrontal dysfunction. 80:754–764. doi:10.1016/j.biopsych.2016....
-
[16]
Diana Rodrigues, Cátia Santa, Bruno Manadas, and Patrícia Monteiro. Chronic stress alters synaptic in- hibition/excitation balance of pyramidal neurons but not PV interneurons in the infralimbic and prelim- bic cortices of C57BL/6J mice. 11:ENEURO.0053–24.2024. doi:10.1523/ENEURO.0053-24.2024. URL http://dx.doi.org/10.1523/ENEURO.0053-24.2024
-
[17]
Eunice Y Yuen, Jing Wei, Wenhua Liu, Ping Zhong, Xiangning Li, and Zhen Yan. Repeated stress causes cognitive impairment by suppressing glutamate receptor expression and function in prefrontal cortex. 73:962–977. doi:10.1016/j.neuron.2011.12.033. URLhttp://dx.doi.org/10.1016/j.neuron.2011.12.033
-
[18]
Circuit- and laminar-specific regulation of medial prefrontal neurons by chronic stress
Wei-Zhu Liu, Chun-Yan Wang, Yu Wang, Mei-Ting Cai, Wei-Xiang Zhong, Tian Liu, Zhi-Hao Wang, Han- Qing Pan, Wen-Hua Zhang, and Bing-Xing Pan. Circuit- and laminar-specific regulation of medial prefrontal neurons by chronic stress. 13:90. doi:10.1186/s13578-023-01050-2. URL http://dx.doi.org/10.1186/ s13578-023-01050-2
-
[19]
Nawshaba Nawreen, Kristen Oshima, James Chambers, Marissa Smail, and James P Herman. Inhibition of prefrontal cortex parvalbumin interneurons mitigates behavioral and physiological sequelae of chronic stress in male mice. 27:2361238. doi:10.1080/10253890.2024.2361238. URL http://dx.doi.org/10.1080/ 10253890.2024.2361238. 19 Chronic stress as an E/I pertur...
-
[20]
Effects of altered excitation-inhibition balance on decision making in a cortical circuit model
Norman H Lam, Thiago Borduqui, Jaime Hallak, Antonio Roque, Alan Anticevic, John H Krystal, Xiao-Jing Wang, and John D Murray. Effects of altered excitation-inhibition balance on decision making in a cortical circuit model. 42:1035–1053. doi:10.1523/JNEUROSCI.1371-20.2021. URL http://dx.doi.org/10.1523/ JNEUROSCI.1371-20.2021
-
[21]
Choice selective inhibition drives stability and competition in decision circuits
James P Roach, Anne K Churchland, and Tatiana A Engel. Choice selective inhibition drives stability and competition in decision circuits. 14:147. doi:10.1038/s41467-023-35822-8. URL http://dx.doi.org/10. 1038/s41467-023-35822-8
-
[22]
H Francis Song, Guangyu R Yang, and Xiao-Jing Wang. Training excitatory-inhibitory recurrent neural networks for cognitive tasks: A simple and flexible framework. 12:e1004792. doi:10.1371/journal.pcbi.1004792. URL http://dx.doi.org/10.1371/journal.pcbi.1004792
-
[23]
Training dynamically balanced excitatory-inhibitory networks
Alessandro Ingrosso and L F Abbott. Training dynamically balanced excitatory-inhibitory networks. 14:e0220547. doi:10.1371/journal.pone.0220547. URLhttp://dx.doi.org/10.1371/journal.pone.0220547
-
[25]
Christopher J Cueva, Adel Ardalan, Misha Tsodyks, and Ning Qian. Recurrent neural network models for working memory of continuous variables: activity manifolds, connectivity patterns, and dynamic codes. doi:10.48550/arXiv.2111.01275. URLhttp://arxiv.org/abs/2111.01275
-
[26]
A recurrent neural network model of prefrontal brain activity during a working memory task
Emilia P Piwek, Mark G Stokes, and Christopher Summerfield. A recurrent neural network model of prefrontal brain activity during a working memory task. 19:e1011555. doi:10.1371/journal.pcbi.1011555. URL http: //dx.doi.org/10.1371/journal.pcbi.1011555
-
[27]
Chengcheng Du, Yinqian Sun, Jihang Wang, Qian Zhang, and Yi Zeng. Synapses mediate the effects of different types of stress on working memory: a brain-inspired spiking neural network study. 19:1534839. doi:10.3389/fncel.2025.1534839. URLhttp://dx.doi.org/10.3389/fncel.2025.1534839
-
[28]
Neuromodulators generate multiple context-relevant behaviors in recurrent neural networks
Ben Tsuda, Stefan C Pate, Kay M Tye, Hava T Siegelmann, and Terrence J Sejnowski. Neuromodulators generate multiple context-relevant behaviors in recurrent neural networks. 38:292–327. doi:10.1162/NECO.a.1489. URL https://dx.doi.org/10.1162/NECO.a.1489
-
[29]
Noisy recur- rent neural networks
Soon Hoe Lim, N Benjamin Erichson, Liam Hodgkinson, and Michael W Mahoney. Noisy recur- rent neural networks. 34:5124–5137. URL https://proceedings.neurips.cc/paper/2021/hash/ 29301521774ff3cbd26652b2d5c95996-Abstract.html
2021
-
[30]
Adversarial weight perturbation helps robust generalization
Dongxian Wu, Shu-Tao Xia, and Yisen Wang. Adversarial weight perturbation helps robust generalization. doi:10.5555/3495724.3495973. URLhttp://dx.doi.org/10.5555/3495724.3495973
-
[31]
Nuttida Rungratsameetaweemana, Robert Kim, Thiparat Chotibut, and Terrence J Sejnowski. Random noise promotes slow heterogeneous synaptic dynamics important for robust working memory computation. 122: e2316745122. doi:10.1073/pnas.2316745122. URLhttp://dx.doi.org/10.1073/pnas.2316745122
-
[32]
Cecilia Jarne and Mariano Caruso. Effect in the spectra of eigenvalues and dynamics of RNNs trained with excitatory-inhibitory constraint. 18:1323–1335. doi:10.1007/s11571-023-09956-w. URL http://dx.doi.org/ 10.1007/s11571-023-09956-w
-
[33]
Functional implications of dale’s law in balanced neuronal network dynamics and decision making
Victor J Barranca, Asha Bhuiyan, Max Sundgren, and Fangzhou Xing. Functional implications of dale’s law in balanced neuronal network dynamics and decision making. 16:801847. doi:10.3389/fnins.2022.801847. URL http://dx.doi.org/10.3389/fnins.2022.801847
-
[34]
Task representations in neural networks trained to perform many cognitive tasks
Guangyu Robert Yang, Madhura R Joglekar, H Francis Song, William T Newsome, and Xiao-Jing Wang. Task representations in neural networks trained to perform many cognitive tasks. 22:297–306. doi:10.1038/s41593- 018-0310-2. URLhttp://dx.doi.org/10.1038/s41593-018-0310-2
-
[35]
Artificial neural networks for neuroscientists: A primer
Guangyu Robert Yang and Xiao-Jing Wang. Artificial neural networks for neuroscientists: A primer. 107:1048–
-
[36]
URLhttp://dx.doi.org/10.1016/j.neuron.2020.09.005
doi:10.1016/j.neuron.2020.09.005. URLhttp://dx.doi.org/10.1016/j.neuron.2020.09.005
-
[37]
Synaptic basis of cortical persistent activity: the importance of NMDA receptors to working memory
X J Wang. Synaptic basis of cortical persistent activity: the importance of NMDA receptors to working memory. 19:9587–9603. doi:10.1523/jneurosci.19-21-09587.1999. URL http://dx.doi.org/10.1523/JNEUROSCI. 19-21-09587.1999
-
[38]
A Compte, N Brunel, P S Goldman-Rakic, and X J Wang. Synaptic mechanisms and network dynamics underlying spatial working memory in a cortical network model. 10:910–923. doi:10.1093/cercor/10.9.910. URL http://dx.doi.org/10.1093/cercor/10.9.910. 20 Chronic stress as an E/I perturbation in RNNsA PREPRINT
-
[39]
Raffael Kalisch, Scott J. Russo, and Marianne B. Müller. Neurobiology and systems biology of stress resilience. Physiological Reviews, 104(3):1205–1263, 2024. doi:10.1152/physrev.00042.2023
-
[40]
Neurobiological basis of stress resilience
Eric J Nestler and Scott J Russo. Neurobiological basis of stress resilience. 112:1911–1929. doi:10.1016/j.neuron.2024.05.001. URLhttp://dx.doi.org/10.1016/j.neuron.2024.05.001
-
[41]
Alfred Rajakumar, John Rinzel, and Zhe S Chen. Stimulus-driven and spontaneous dynamics in excitatory- inhibitory recurrent neural networks for sequence representation. 33:2603–2645. doi:10.1162/neco_a_01418. URLhttp://dx.doi.org/10.1162/neco_a_01418
-
[42]
Predictive coding is a consequence of energy efficiency in recurrent neural networks
Abdullahi Ali, Nasir Ahmad, Elgar de Groot, Marcel Antonius Johannes van Gerven, and Tim Christian Ki- etzmann. Predictive coding is a consequence of energy efficiency in recurrent neural networks. 3:100639. doi:10.1016/j.patter.2022.100639. URLhttp://dx.doi.org/10.1016/j.patter.2022.100639
-
[43]
David Sussillo and Omri Barak. Opening the black box: low-dimensional dynamics in high-dimensional recurrent neural networks. 25:626–649. doi:10.1162/NECO_a_00409. URL http://dx.doi.org/10.1162/NECO_a_ 00409
-
[44]
Dynamical system approach to explainability in recurrent neural networks
Alexis Dubreuil. Dynamical system approach to explainability in recurrent neural networks. URL https://www. semanticscholar.org/paper/Dynamical-system-approach-to-explainability-in-Dubreuil/ d08515aefa1a9df330a33106e788e21daa14c135
-
[45]
Considerations in using recurrent neural networks to probe neural dynamics
Jonathan C Kao. Considerations in using recurrent neural networks to probe neural dynamics. 122:2504–2521. doi:10.1152/jn.00467.2018. URLhttp://dx.doi.org/10.1152/jn.00467.2018
-
[46]
A neural network walks into a lab: towards using deep nets as models for human behavior
Wei Ji Ma and Benjamin Peters. A neural network walks into a lab: towards using deep nets as models for human behavior. doi:10.48550/arXiv.2005.02181. URLhttp://arxiv.org/abs/2005.02181
-
[47]
General principles of neuronal co-transmission: Insights from multiple model systems
Erik Svensson, John Apergis-Schoute, Geoffrey Burnstock, Michael P Nusbaum, David Parker, and Helgi B Schiöth. General principles of neuronal co-transmission: Insights from multiple model systems. 12:117. doi:10.3389/fncir.2018.00117. URLhttp://dx.doi.org/10.3389/fncir.2018.00117
-
[48]
Marian Joëls and Tallie Z Baram. The neuro-symphony of stress. 10:459–466. doi:10.1038/nrn2632. URL http://dx.doi.org/10.1038/nrn2632
-
[49]
The neocortical circuit: themes and variations
Kenneth D Harris and Gordon M G Shepherd. The neocortical circuit: themes and variations. 18:170–181. doi:10.1038/nn.3917. URLhttp://dx.doi.org/10.1038/nn.3917
-
[50]
P Strata and R Harvey. Dale’s principle. 50:349–350. doi:10.1016/s0361-9230(99)00100-8. URL http: //dx.doi.org/10.1016/s0361-9230(99)00100-8
-
[51]
Neuronal correlates of parametric working memory in the prefrontal cortex
R Romo, C D Brody, A Hernández, and L Lemus. Neuronal correlates of parametric working memory in the prefrontal cortex. 399:470–473. doi:10.1038/20939. URLhttp://dx.doi.org/10.1038/20939
-
[52]
Neuronal population coding of parametric working memory
Omri Barak, Misha Tsodyks, and Ranulfo Romo. Neuronal population coding of parametric working memory. 30:9424–9430. doi:10.1523/JNEUROSCI.1875-10.2010. URL http://dx.doi.org/10.1523/JNEUROSCI. 1875-10.2010
-
[53]
JAX: composable transformations of Python+NumPy programs
James Bradbury, Roy Frostig, Peter Hawkins, Matthew James Johnson, Yash Katariya, Chris Leary, Dougal Maclaurin, George Necula, Adam Paszke, Jake VanderPlas, Skye Wanderman-Milne, and Qiao Zhang. JAX: composable transformations of Python+NumPy programs. URLhttp://github.com/jax-ml/jax
-
[54]
Charles R Harris, K Jarrod Millman, Stéfan J van der Walt, Ralf Gommers, Pauli Virtanen, David Cournapeau, Eric Wieser, Julian Taylor, Sebastian Berg, Nathaniel J Smith, Robert Kern, Matti Picus, Stephan Hoyer, Marten H van Kerkwijk, Matthew Brett, Allan Haldane, Jaime Fernández del Río, Mark Wiebe, Pearu Peterson, Pierre Gérard-Marchant, Kevin Sheppard, ...
-
[55]
SciPy1.0: FundamentalalgorithmsforscientificcomputinginPython,
Pauli Virtanen, Ralf Gommers, Travis E Oliphant, Matt Haberland, Tyler Reddy, David Cournapeau, Evgeni Burovski, Pearu Peterson, Warren Weckesser, Jonathan Bright, Stéfan J van der Walt, Matthew Brett, Joshua Wilson, K Jarrod Millman, Nikolay Mayorov, Andrew R J Nelson, Eric Jones, Robert Kern, Eric Larson, C J Carey, ˙Ilhan Polat, Yu Feng, Eric W Moore, ...
-
[56]
Data Structures for Statistical Computing in Python
Wes McKinney. Data structures for statistical computing in python. In Stéfan van der Walt and Jarrod Millman, editors,Proceedings of the 9th Python in Science Conference, pages 56–61. doi:10.25080/Majora-92bf1922-00a. URLhttp://dx.doi.org/10.25080/Majora-92bf1922-00a. 21 Chronic stress as an E/I perturbation in RNNsA PREPRINT
-
[57]
Scikit-learn: Machine learning in python
F Pedregosa, G Varoquaux, A Gramfort, V Michel, B Thirion, O Grisel, M Blondel, P Prettenhofer, R Weiss, V Dubourg, J Vanderplas, A Passos, D Cournapeau, M Brucher, M Perrot, and E Duchesnay. Scikit-learn: Machine learning in python. 12:2825–2830
-
[58]
statsmodels: Econometric and statistical modeling with python
Skipper Seabold and Josef Perktold. statsmodels: Econometric and statistical modeling with python. In9th Python in Science Conference
-
[59]
Matplotlib: A 2D Graphics Environment
J D Hunter. Matplotlib: A 2D graphics environment. 9:90–95. doi:10.1109/MCSE.2007.55. URL http: //dx.doi.org/10.1109/MCSE.2007.55. 22
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.