FLYNN: Robust Neural Network for Robot Navigation using Fly Brain Topology
Pith reviewed 2026-07-02 21:49 UTC · model grok-4.3
The pith
A neural network built from the fruit fly brain connectome navigates robots with greater tolerance to sensory loss than standard networks.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
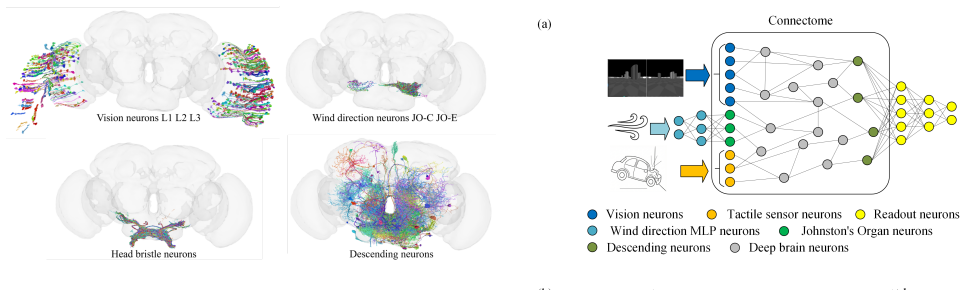
The fly connectome neural network (FLYNN) achieves performance comparable to modern hand-crafted networks in MuJoCo vision-based navigation while exhibiting superior resistance to out-of-distribution data and tolerance to sensory loss without further training. It remained functional even under total vision loss while hand-crafted networks largely failed, even when specifically trained with camera dropout. Principal Component Analysis of the internal state suggests a high degree of representational modularity related to its robustness.
What carries the argument
FLYNN, the recurrent neural network whose architecture is directly derived from the synaptic-resolution brain connectome of Drosophila melanogaster.
If this is right
- FLYNN maintains function under total vision loss without any retraining.
- It shows greater resistance to out-of-distribution inputs than hand-crafted networks of similar size.
- Representational modularity observed via PCA may underlie the robustness advantage.
- Using biological brain topology offers a direction for building resilient artificial agents.
Where Pith is reading between the lines
- The connectome mapping approach could be applied to other robot control problems such as grasping or locomotion.
- Running the trained network on physical hardware would test whether the simulated robustness carries over to real sensors and motors.
- The observed modularity could guide construction of mixed networks that blend fly wiring with other topologies.
- If the robustness stems from evolutionary tuning of the connectome, networks derived from other organisms might show similar properties.
Load-bearing premise
The fly brain's synaptic wiring pattern, when copied into an RNN structure and trained on navigation, will automatically transfer the biological network's tolerance to missing sensors and new conditions.
What would settle it
Testing FLYNN and the hand-crafted networks on the same navigation task but in a different simulated environment or with additional sensor noise types and finding that the performance advantage disappears.
Figures






read the original abstract
While deep learning models achieve state-of-the-art performance in complex tasks, they remain brittle when faced with new environments or sensory deprivation. In contrast, biological systems exhibit remarkable tolerance to these challenges. We address this vulnerability by developing a recurrent neural network (RNN) whose architecture is directly derived from the synaptic-resolution brain connectome of the fruit fly Drosophila melanogaster. We demonstrate the feasibility of training the fly connectome neural network (FLYNN) to perform vision-based navigation in MuJoCo, achieving performance comparable to modern hand-crafted networks of similar parameter counts. Crucially, FLYNN exhibits superior resistance to out-of-distribution (OOD) data and tolerance to sensory loss without further training. It remained functional even under total vision loss while hand-crafted networks largely failed, even when specifically trained with camera dropout. Principal Component Analysis (PCA) of the internal state of FLYNN suggests that it exhibits a particularly high degree of representational modularity, which might be related to its robustness. Our work provides a new direction for designing resilient artificial agents following the topology of biological brains.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces FLYNN, a recurrent neural network whose architecture is directly derived from the synaptic-resolution Drosophila melanogaster connectome. The network is trained to perform vision-based navigation in MuJoCo and is reported to achieve performance comparable to hand-crafted networks of similar size while exhibiting superior robustness to out-of-distribution inputs and to sensory loss (including total vision loss) without retraining. PCA of internal states is used to suggest higher representational modularity as a possible mechanism for the observed robustness.
Significance. If the central claim holds after appropriate controls, the work would provide concrete evidence that biological connectome topologies can be mapped to artificial networks to confer robustness properties not easily obtained from standard hand-crafted architectures. This would constitute a useful empirical bridge between neuroscience and robotics, with potential implications for designing agents tolerant to sensor failure.
major comments (2)
- [Results on robustness experiments] Results section on robustness to sensory loss and OOD inputs: the attribution of superior tolerance (including functionality under total vision loss) to the specific Drosophila connectivity pattern is not supported by the presented evidence. The experiments compare only against hand-crafted networks; no null-model controls (e.g., degree-sequence-preserving edge shuffles or other randomized topologies with matched statistics) are reported. Without these, it remains possible that the robustness arises from generic RNN properties, training procedure, or parameter count rather than the biological topology itself.
- [Methods / Network architecture] Section describing the network construction and training: the mapping from the fly connectome to the RNN is presented as direct, yet no quantitative details on how synaptic weights, neuron types, or recurrent dynamics are instantiated from the connectome data are supplied, nor are ablation studies on alternative mappings provided. This makes it difficult to assess whether the claimed robustness is reproducible or tied to the specific biological topology.
minor comments (2)
- [Abstract] The abstract states performance and robustness results without any numerical values, error bars, or statistical tests; these should be added for clarity.
- [Discussion] The PCA modularity analysis is presented as suggestive but post-hoc; its relation to the robustness results would benefit from a more quantitative link (e.g., correlation with robustness metrics across conditions).
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive comments. We address each major point below and outline revisions that will strengthen the attribution of robustness to the biological topology and improve reproducibility of the network construction.
read point-by-point responses
-
Referee: [Results on robustness experiments] Results section on robustness to sensory loss and OOD inputs: the attribution of superior tolerance (including functionality under total vision loss) to the specific Drosophila connectivity pattern is not supported by the presented evidence. The experiments compare only against hand-crafted networks; no null-model controls (e.g., degree-sequence-preserving edge shuffles or other randomized topologies with matched statistics) are reported. Without these, it remains possible that the robustness arises from generic RNN properties, training procedure, or parameter count rather than the biological topology itself.
Authors: We agree that null-model controls are necessary to isolate the contribution of the specific Drosophila connectivity. In the revised manuscript we will add experiments using degree-sequence-preserving edge shuffles and other topology-preserving randomizations with matched statistics (e.g., clustering coefficient, motif counts). These controls will be trained and evaluated under identical conditions to demonstrate that the observed robustness exceeds what is expected from generic RNN properties or parameter count alone. revision: yes
-
Referee: [Methods / Network architecture] Section describing the network construction and training: the mapping from the fly connectome to the RNN is presented as direct, yet no quantitative details on how synaptic weights, neuron types, or recurrent dynamics are instantiated from the connectome data are supplied, nor are ablation studies on alternative mappings provided. This makes it difficult to assess whether the claimed robustness is reproducible or tied to the specific biological topology.
Authors: We will substantially expand the Methods section with quantitative details: synaptic counts and weights from the connectome will be linearly scaled to initialize RNN recurrent weights within a defined range; neuron types (e.g., cholinergic, GABAergic) will be mapped to specific activation functions based on published physiological data; and recurrent dynamics will be instantiated via the exact adjacency matrix with self-connections removed. We will also add ablation experiments that (i) randomize weights while preserving topology and (ii) use alternative mappings (e.g., uniform random initialization of the same graph). These additions will allow readers to assess reproducibility and the role of the biological topology. revision: yes
Circularity Check
No significant circularity; robustness claim is empirical outcome
full rationale
The paper maps the Drosophila connectome to an RNN architecture, trains it on MuJoCo navigation, and reports empirical robustness results (OOD tolerance, sensory loss) versus hand-crafted baselines. No derivation step reduces a claimed result to its inputs by construction: there are no fitted parameters renamed as predictions, no self-citation chains justifying uniqueness of the topology, and no ansatz or renaming that collapses the central claim. The PCA modularity observation is post-hoc and does not bear on the training or robustness metrics. The derivation chain remains independent of the reported performance numbers.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The Drosophila connectome can be faithfully converted into an RNN weight matrix without loss of functional properties relevant to robustness.
Reference graph
Works this paper leans on
-
[1]
The perceptron: a probabilistic model for information storage and organization in the brain
F. Rosenblatt, “The perceptron: a probabilistic model for information storage and organization in the brain.”Psychological review, vol. 65, no. 6, p. 386, 1958
work page 1958
-
[2]
Handwritten digit recognition with a back-propagation network,
Y . LeCun, B. Boser, J. Denker, D. Henderson, R. Howard, W. Hubbard, and L. Jackel, “Handwritten digit recognition with a back-propagation network,” inAdvances in Neural Information Processing Systems, D. Touretzky, Ed., vol. 2. Morgan-Kaufmann, 1989
work page 1989
-
[3]
Imagenet classification with deep convolutional neural networks,
A. Krizhevsky, I. Sutskever, and G. E. Hinton, “Imagenet classification with deep convolutional neural networks,” inAdvances in Neural Information Processing Systems, F. Pereira, C. Burges, L. Bottou, and K. Weinberger, Eds., vol. 25. Curran Associates, Inc., 2012
work page 2012
-
[4]
Backpropagation through time: what it does and how to do it,
P. J. Werbos, “Backpropagation through time: what it does and how to do it,”Proceedings of the IEEE, vol. 78, no. 10, pp. 1550–1560, 2002
work page 2002
-
[5]
S. Hochreiter and J. Schmidhuber, “Long short-term memory,”Neural computation, vol. 9, no. 8, pp. 1735–1780, 1997
work page 1997
-
[6]
A. Vaswani, N. Shazeer, N. Parmar, J. Uszkoreit, L. Jones, A. N. Gomez, Ł. Kaiser, and I. Polosukhin, “Attention is all you need,” Advances in neural information processing systems, vol. 30, 2017
work page 2017
-
[7]
Out-of-distribution seg- mentation in autonomous driving: Problems and state of the art,
Y . Shoeb, A. Nowzad, and H. Gottschalk, “Out-of-distribution seg- mentation in autonomous driving: Problems and state of the art,” inProceedings of the Computer Vision and Pattern Recognition Conference, 2025, pp. 4310–4320
work page 2025
-
[8]
Rgb cameras failures and their effects in autonomous driving applications,
A. Ceccarelli and F. Secci, “Rgb cameras failures and their effects in autonomous driving applications,”IEEE Transactions on Dependable and Secure Computing, vol. 20, no. 4, pp. 2731–2745, 2022
work page 2022
-
[9]
The drosophila visual system: From neural circuits to behavior,
Y . Zhu, “The drosophila visual system: From neural circuits to behavior,”Cell adhesion & migration, vol. 7, no. 4, pp. 333–344, 2013
work page 2013
-
[10]
A. Borst and L. N. Groschner, “How flies see motion,”Annual review of neuroscience, vol. 46, no. 1, pp. 17–37, 2023
work page 2023
-
[11]
The neuroanatomical ultrastructure and function of a biological ring attractor,
D. B. Turner-Evans, K. T. Jensen, S. Ali, T. Paterson, A. Sheridan, R. P. Ray, T. Wolff, J. S. Lauritzen, G. M. Rubin, D. D. Bock,et al., “The neuroanatomical ultrastructure and function of a biological ring attractor,”Neuron, vol. 108, no. 1, pp. 145–163, 2020
work page 2020
-
[12]
Flexible navigational computations in the drosophila central complex,
Y . E. Fisher, “Flexible navigational computations in the drosophila central complex,”Current opinion in neurobiology, vol. 73, p. 102514, 2022
work page 2022
-
[13]
Mobile robot navigation via domain randomization and real-world adaptation,
P. Qin, T. Zhao, Q. Su, and H. Yang, “Mobile robot navigation via domain randomization and real-world adaptation,” in2025 IEEE International Conference on Unmanned Systems (ICUS). IEEE, 2025, pp. 878–883
work page 2025
-
[14]
Robot vision-based autonomous navigation method using sim2real domain adaptation,
M. Zhang, S. Duan, and C. Xu, “Robot vision-based autonomous navigation method using sim2real domain adaptation,” in2024 IEEE International Symposium on Parallel and Distributed Processing with Applications (ISPA). IEEE, 2024, pp. 1203–1209
work page 2024
-
[15]
Liquid neural networks for autonomous driving: A framework for intelligent decision-making,
Y . Suresh, V . Devashya,et al., “Liquid neural networks for autonomous driving: A framework for intelligent decision-making,” in2025 5th International Conference on Emerging Research in Electronics, Com- puter Science and Technology (ICERECT). IEEE, 2025, pp. 1–6
work page 2025
-
[16]
Robust flight navigation out of distribution with liquid neural networks,
M. Chahine, R. Hasani, P. Kao, A. Ray, R. Shubert, M. Lechner, A. Amini, and D. Rus, “Robust flight navigation out of distribution with liquid neural networks,”Science Robotics, vol. 8, no. 77, p. eadc8892, 2023
work page 2023
-
[17]
Generating out-of-distribution scenarios using language models,
E. Aasi, P. Nguyen, S. Sreeram, G. Rosman, S. Karaman, and D. Rus, “Generating out-of-distribution scenarios using language models,” in 2025 IEEE International Conference on Robotics and Automation (ICRA). IEEE, 2025, pp. 10 616–10 623
work page 2025
-
[18]
Y . Qiao, W. Lyu, H. Wang, Z. Wang, Z. Li, Y . Zhang, M. Tan, and Q. Wu, “Open-nav: Exploring zero-shot vision-and-language naviga- tion in continuous environment with open-source llms,” in2025 IEEE International Conference on Robotics and Automation (ICRA). IEEE, 2025, pp. 6710–6717
work page 2025
-
[19]
Misconv: Convolutional neural networks for missing data,
M. Przewiezlikowski, M. Smieja, L. Struski, and J. Tabor, “Misconv: Convolutional neural networks for missing data,” inProceedings of the IEEE/CVF Winter Conference on Applications of Computer Vision (WACV), January 2022, pp. 2060–2069
work page 2022
-
[20]
Are deep learning models robust to partial object occlusion in visual recognition tasks,
K. Kassaw, F. Luzi, L. M. Collins, and J. M. Malof, “Are deep learning models robust to partial object occlusion in visual recognition tasks,” Pattern Recognition, vol. 171, p. 112215, 2026
work page 2026
-
[21]
A deep learning gated architecture for ugv navigation robust to sensor failures,
N. Patel, A. Choromanska, P. Krishnamurthy, and F. Khorrami, “A deep learning gated architecture for ugv navigation robust to sensor failures,”Robotics and Autonomous Systems, vol. 116, pp. 80–97, 2019
work page 2019
-
[22]
Neuronal wiring diagram of an adult brain,
S. Dorkenwald, A. Matsliah, A. R. Sterling, P. Schlegel, S.-C. Yu, C. E. McKellar, A. Lin, M. Costa, K. Eichler, Y . Yin,et al., “Neuronal wiring diagram of an adult brain,”Nature, vol. 634, no. 8032, pp. 124– 138, 2024
work page 2024
-
[23]
Whole- brain annotation and multi-connectome cell typing of drosophila,
P. Schlegel, Y . Yin, A. S. Bates, S. Dorkenwald, K. Eichler, P. Brooks, D. S. Han, M. Gkantia, M. Dos Santos, E. J. Munnelly,et al., “Whole- brain annotation and multi-connectome cell typing of drosophila,” Nature, vol. 634, no. 8032, pp. 139–152, 2024
work page 2024
-
[24]
Connectome-constrained networks predict neural activity across the fly visual system,
J. K. Lappalainen, F. D. Tschopp, S. Prakhya, M. McGill, A. Nern, K. Shinomiya, S.-y. Takemura, E. Gruntman, J. H. Macke, and S. C. Turaga, “Connectome-constrained networks predict neural activity across the fly visual system,”Nature, vol. 634, no. 8036, pp. 1132– 1140, 2024
work page 2024
-
[25]
A drosophila computational brain model reveals sensorimotor processing,
P. K. Shiu, G. R. Sterne, N. Spiller, R. Franconville, A. Sandoval, J. Zhou, N. Simha, C. H. Kang, S. Yu, J. S. Kim,et al., “A drosophila computational brain model reveals sensorimotor processing,”Nature, vol. 634, no. 8032, pp. 210–219, 2024. [Online]. Available: https://www.nature.com/articles/s41586-024-07763-9
work page 2024
-
[26]
Connectomic analysis of taste circuits in drosophila,
S. R. Walker, M. Pe ˜na-Garcia, and A. V . Devineni, “Connectomic analysis of taste circuits in drosophila,”Scientific Reports, vol. 15, no. 1, p. 5278, 2025
work page 2025
-
[27]
Mujoco: A physics engine for model-based control,
E. Todorov, T. Erez, and Y . Tassa, “Mujoco: A physics engine for model-based control,” in2012 IEEE/RSJ International Conference on Intelligent Robots and Systems, 2012, pp. 5026–5033
work page 2012
-
[28]
EfficientNet: Rethinking model scaling for convolutional neural networks,
M. Tan and Q. Le, “EfficientNet: Rethinking model scaling for convolutional neural networks,” inProceedings of the 36th Interna- tional Conference on Machine Learning, ser. Proceedings of Machine Learning Research, K. Chaudhuri and R. Salakhutdinov, Eds., vol. 97. PMLR, 09–15 Jun 2019, pp. 6105–6114
work page 2019
-
[29]
A. Howard, M. Sandler, G. Chu, L.-C. Chen, B. Chen, M. Tan, W. Wang, Y . Zhu, R. Pang, V . Vasudevan, Q. V . Le, and H. Adam, “Searching for mobilenetv3,” inProceedings of the IEEE/CVF Inter- national Conference on Computer Vision (ICCV), October 2019
work page 2019
-
[30]
Collective dynamics of ‘small- world’networks,
D. J. Watts and S. H. Strogatz, “Collective dynamics of ‘small- world’networks,”nature, vol. 393, no. 6684, pp. 440–442, 1998
work page 1998
-
[31]
H. Kalmus, “Anemotaxis in drosophila,”Nature, vol. 150, no. 3805, pp. 405–405, 1942
work page 1942
-
[32]
The drosophila auditory system,
G. Boekhoff-Falk and D. F. Eberl, “The drosophila auditory system,” Wiley Interdisciplinary Reviews: Developmental Biology, vol. 3, no. 2, pp. 179–191, 2014
work page 2014
-
[33]
A reduction of imitation learning and structured prediction to no-regret online learning,
S. Ross, G. Gordon, and D. Bagnell, “A reduction of imitation learning and structured prediction to no-regret online learning,” inProceedings of the fourteenth international conference on artificial intelligence and statistics. JMLR Workshop and Conference Proceedings, 2011, pp. 627–635
work page 2011
-
[34]
Vfh*: local obstacle avoidance with look- ahead verification,
I. Ulrich and J. Borenstein, “Vfh*: local obstacle avoidance with look- ahead verification,” inProceedings 2000 ICRA. Millennium Confer- ence. IEEE International Conference on Robotics and Automation. Symposia Proceedings (Cat. No.00CH37065), vol. 3, 2000, pp. 2505– 2511 vol.3
work page 2000
-
[35]
Small-world brain networks revisited,
D. S. Bassett and E. T. Bullmore, “Small-world brain networks revisited,”The Neuroscientist, vol. 23, no. 5, pp. 499–516, 2017
work page 2017
-
[36]
Exploring randomly wired neural networks for image recognition,
S. Xie, A. Kirillov, R. Girshick, and K. He, “Exploring randomly wired neural networks for image recognition,” inProceedings of the IEEE/CVF international conference on computer vision, 2019, pp. 1284–1293
work page 2019
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.