Modeling tumor cell heterogeneity and plasticity in adaptive therapy
Pith reviewed 2026-05-19 23:20 UTC · model grok-4.3
The pith
A mathematical model with continuous drug susceptibility shows adaptive therapy controls tumors longer than fixed dosing by regulating sensitive cell populations.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
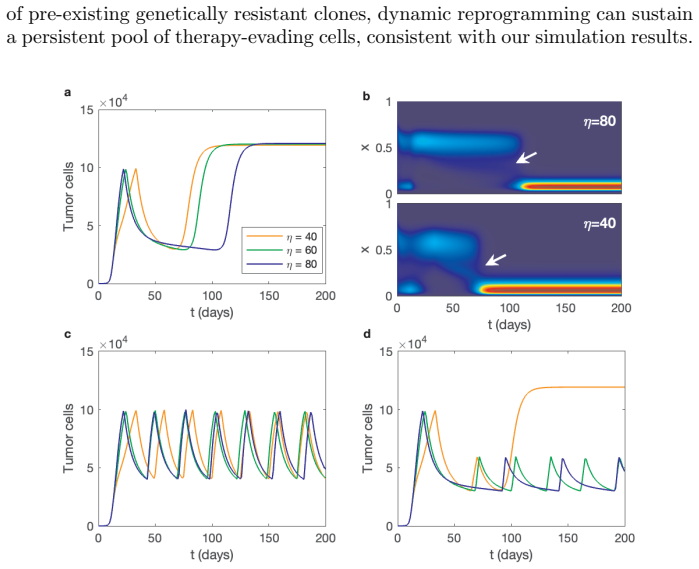
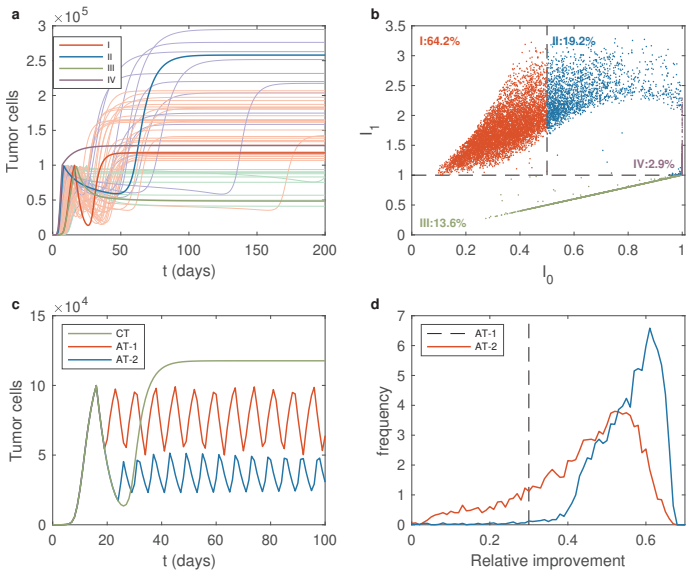
The proposed integro-differential system integrates a continuous drug susceptibility index with a probabilistic inheritance function to describe clonal dynamics under therapy. The resulting model generalizes traditional two-type competition models and captures both heterogeneity and plasticity of tumor cells. Analytical and numerical studies show that continuous therapy drives rapid expansion of resistant clones, adaptive therapy maintains long-term tumor control by dynamically regulating sensitive populations, and high phenotypic plasticity accelerates phenotype switching, leading to earlier tumor relapse following continuous therapy.
What carries the argument
The integro-differential system built from a continuous drug susceptibility index and a probabilistic inheritance function, which integrates over susceptibility levels to track how heterogeneity and plasticity shape clonal competition under different treatment schedules.
If this is right
- Continuous therapy selects for resistant clones faster than adaptive schedules.
- Adaptive therapy sustains control by keeping sensitive populations active in competition.
- Elevated phenotypic plasticity shortens time to relapse after continuous treatment ends.
- Specific ranges of susceptibility and inheritance parameters favor adaptive over fixed regimens.
Where Pith is reading between the lines
- The framework could be used to test whether measuring a patient's tumor susceptibility distribution would improve schedule design.
- Similar continuous-state models might apply to resistance management in bacterial infections or viral populations.
- Spatial or immune effects could be added to see whether they amplify or dampen the reported plasticity-driven relapse differences.
Load-bearing premise
The probabilistic inheritance function and continuous drug susceptibility index are assumed to sufficiently represent real biological plasticity and heterogeneity in tumor cells without specific empirical calibration or validation data.
What would settle it
Compare measured relapse times and phenotype-switching rates in tumor cell lines or mouse models treated with continuous versus adaptive regimens; if high plasticity fails to produce earlier relapse under continuous therapy, the model's central predictions do not hold.
Figures






read the original abstract
Adaptive therapy (AT) is designed to postpone the emergence of drug resistance by exploiting evolutionary competition among tumor subclones. Most mathematical models of AT assume a binary population structure of drug-sensitive and drug-resistant cells, which neglects the continuous nature of phenotypic plasticity. In this study, we propose a mathematical model that integrates a continuous drug susceptibility index with a probabilistic inheritance function to describe clonal dynamics under therapy. The resulting integro-differential system generalizes traditional two-type competition models and captures both heterogeneity and plasticity of tumor cells. Analytical and numerical studies show that (i) continuous therapy drives rapid expansion of resistant clones, (ii) adaptive therapy maintains long-term tumor control by dynamically regulating sensitive populations, and (iii) high phenotypic plasticity accelerates phenotype switching, leading to earlier tumor relapse following continuous therapy. These results identify critical parameter regimes where adaptive therapy outperforms fixed regimens and highlight the essential role of plasticity in shaping treatment outcomes. The proposed framework provides a more realistic mathematical foundation for the design of clinically relevant adaptive therapy strategies.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops an integro-differential model that augments traditional two-type competition frameworks with a continuous drug susceptibility index and a probabilistic inheritance function for phenotype switching. Analytical and numerical analyses are used to argue that (i) continuous therapy accelerates expansion of resistant clones, (ii) adaptive therapy achieves sustained tumor control by dynamically suppressing sensitive subpopulations, and (iii) elevated phenotypic plasticity hastens switching and precipitates earlier relapse under continuous regimens, thereby delineating parameter regimes favoring adaptive strategies.
Significance. If the central derivations and simulations are robust, the work supplies a mathematically consistent extension of binary models that explicitly incorporates continuous heterogeneity and plasticity, offering a clearer mechanistic basis for why adaptive therapy can outperform fixed dosing and for the sensitivity of outcomes to switching rates.
major comments (2)
- [Model formulation] Model section (definition of the probabilistic inheritance function and susceptibility index): the qualitative claims (i)–(iii) rest on the specific functional forms chosen for these kernels; no calibration to measured switching rates, clonal frequency distributions, or drug-response curves is provided, so the reported superiority of adaptive therapy and the plasticity-relapse link remain conditional on untested assumptions about memoryless switching and susceptibility gradients.
- [Numerical studies] Numerical results (parameter sweeps and regime diagrams): the manuscript should demonstrate that the reported thresholds for long-term control and earlier relapse are insensitive to modest perturbations of the inheritance kernel parameters; without such checks, the load-bearing distinction between high- and low-plasticity regimes cannot be considered reliable.
minor comments (2)
- [Abstract] Abstract: the statement that 'analytical and numerical studies support' the three findings would be strengthened by a one-sentence indication of the key analytical technique (e.g., stability analysis of the integro-differential system) or the numerical scheme employed.
- [Model formulation] Notation: the continuous susceptibility index is referred to both as a distribution and as a dynamic variable; a single consistent symbol and a brief clarification of its time evolution would remove ambiguity.
Simulated Author's Rebuttal
We thank the referee for their detailed and constructive comments. We address each major comment below and have revised the manuscript to strengthen the presentation of assumptions and to include additional robustness checks.
read point-by-point responses
-
Referee: [Model formulation] Model section (definition of the probabilistic inheritance function and susceptibility index): the qualitative claims (i)–(iii) rest on the specific functional forms chosen for these kernels; no calibration to measured switching rates, clonal frequency distributions, or drug-response curves is provided, so the reported superiority of adaptive therapy and the plasticity-relapse link remain conditional on untested assumptions about memoryless switching and susceptibility gradients.
Authors: We agree that the qualitative results depend on the chosen functional forms for the inheritance kernel and susceptibility index. These forms were selected to capture memoryless switching and continuous gradients in a manner consistent with existing phenotypic plasticity literature, but we acknowledge that direct calibration to specific datasets is not performed here. In the revised manuscript we have added a dedicated paragraph in the Discussion section that (i) motivates the functional choices with references to empirical switching-rate studies, (ii) explicitly states the scope of the claims as exploratory rather than predictive for any particular tumor, and (iii) reports supplementary simulations using alternative kernel shapes (Gaussian versus exponential decay) that preserve the reported ordering of adaptive versus continuous therapy outcomes. These additions clarify the conditional nature of the findings without altering the core model. revision: partial
-
Referee: [Numerical studies] Numerical results (parameter sweeps and regime diagrams): the manuscript should demonstrate that the reported thresholds for long-term control and earlier relapse are insensitive to modest perturbations of the inheritance kernel parameters; without such checks, the load-bearing distinction between high- and low-plasticity regimes cannot be considered reliable.
Authors: We appreciate this suggestion for strengthening the numerical evidence. In the revised version we have added a new supplementary figure (Fig. S3) together with a short subsection in the Numerical Methods that performs local sensitivity analysis on the inheritance-kernel parameters. Specifically, we vary the mean switching rate and the width parameter by ±20 % around the baseline values used in the main text and recompute the critical plasticity thresholds for relapse under continuous therapy. The additional results show that the qualitative separation between high- and low-plasticity regimes, as well as the advantage of adaptive therapy, remains intact under these perturbations. The revised text now explicitly references these checks when discussing the plasticity-relapse link. revision: yes
Circularity Check
No circularity: results follow from independent model analysis
full rationale
The paper defines a new integro-differential model using a continuous drug susceptibility index and probabilistic inheritance function as core ingredients to generalize binary population models. Analytical and numerical studies then derive the listed behaviors (rapid resistant expansion under continuous therapy, long-term control under adaptive therapy, and plasticity effects on relapse timing) directly from the dynamics of this system. No step reduces a claimed prediction to a fitted parameter or self-referential definition by construction, and the abstract contains no load-bearing self-citations or uniqueness theorems imported from prior author work. The derivation chain is therefore self-contained as standard mathematical exploration of an explicitly posited framework.
Axiom & Free-Parameter Ledger
free parameters (2)
- parameters of the probabilistic inheritance function
- parameters defining the continuous drug susceptibility index distribution
axioms (2)
- domain assumption Tumor cells exhibit continuous rather than discrete variation in drug susceptibility.
- domain assumption Phenotypic inheritance can be represented by a probabilistic function that governs clonal dynamics under therapy.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
we propose a mathematical model that integrates a continuous drug susceptibility index with a probabilistic inheritance function... p(x, y) = xa(y)−1(1−x)b(y)−1 / B(a(y), b(y))
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The resulting integro-differential system generalizes traditional two-type competition models
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Siegel, Mathieu Laversanne, Isabelle Soerjo- mataram, Ahmedin Jemal, and Freddie Bray
Hyuna Sung, Jacques Ferlay, Rebecca L. Siegel, Mathieu Laversanne, Isabelle Soerjo- mataram, Ahmedin Jemal, and Freddie Bray. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries.CA: Cancer J Clin, 71(3):209–249, February 2021
work page 2020
-
[2]
Mel Greaves and Carlo C. Maley. Clonal evolution in cancer.Nature, 481(7381):306–313, January 2012
work page 2012
-
[3]
Robert A. Gatenby, Ariosto S. Silva, Robert J. Gillies, and B. Roy Frieden. Adaptive therapy.Cancer Res, 69(11):4894–4903, June 2009
work page 2009
-
[4]
Robert A. Gatenby and Joel S. Brown. Integrating evolutionary dynamics into cancer therapy.Nat Rev Clin Oncol, 17(11):675–686, July 2020
work page 2020
-
[5]
C. Athena Aktipis and Randolph M. Nesse. Evolutionary foundations for cancer biology. Evol Appl, 6(1):144–159, January 2013
work page 2013
-
[6]
Jill A. Gallaher, Pedro M. Enriquez-Navas, Kimberly A. Luddy, Robert A. Gatenby, and Alexander R.A. Anderson. Spatial heterogeneity and evolutionary dynamics modulate time to recurrence in continuous and adaptive cancer therapies.Cancer Res, 78(8):2127– 2139, April 2018
work page 2018
-
[7]
Gillies, Daniel Verduzco, and Robert A
Robert J. Gillies, Daniel Verduzco, and Robert A. Gatenby. Evolutionary dynamics of carcinogenesis and why targeted therapy does not work.Nat Rev Cancer, 12(7):487–493, June 2012
work page 2012
-
[8]
Jingsong Zhang, Jessica J. Cunningham, Joel S. Brown, and Robert A. Gatenby. In- tegrating evolutionary dynamics into treatment of metastatic castrate-resistant prostate cancer.Nat Commun, 8(1), November 2017
work page 2017
-
[9]
Brown, Zeynep Eroglu, and Alexander R.A
Eunjung Kim, Joel S. Brown, Zeynep Eroglu, and Alexander R.A. Anderson. Adap- tive therapy for metastatic melanoma: Predictions from patient calibrated mathematical models.Cancers, 13(4):823, February 2021
work page 2021
-
[10]
Ruiyang Liu, Shun Wang, Xuewen Tan, and Xiufen Zou. Identifying optimal adaptive therapeutic schedules for prostate cancer through combining mathematical modeling and dynamic optimization.Appl Math Model, 107:688–700, July 2022
work page 2022
-
[11]
Jingsong Zhang, Jessica Cunningham, Joel Brown, and Robert Gatenby. Evolution- based mathematical models significantly prolong response to abiraterone in metastatic castrate-resistant prostate cancer and identify strategies to further improve outcomes. eLife, 11:e76284, Jun 2022
work page 2022
-
[12]
Dujuan Wang and Jinzhi Lei. Optimal adaptive therapeutic schedules for metastatic castrate-resistant prostate cancer based on bilevel optimization problem.J Math Biol, 90:60, 2025
work page 2025
-
[13]
Cordelia McGehee and Yoichiro Mori. A mathematical framework for comparison of intermittent versus continuous adaptive chemotherapy dosing in cancer.npj Syst Biol Appl, 10(1):140, November 2024
work page 2024
-
[14]
Yoonseok Kam, Tuhin Das, Susan Minton, and Robert A Gatenby. Evolutionary strategy for systemic therapy of metastatic breast cancer: Balancing response with suppression of resistance.Women ’s Health(Lond), 10(4):423–430, July 2014
work page 2014
-
[15]
Jana L. Gevertz, James M. Greene, Samantha Prosperi, Natacha Comandante-Lou, and Eduardo D. Sontag. Understanding therapeutic tolerance through a mathematical model of drug-induced resistance.npj Syst Biol Appl, 11(1), April 2025
work page 2025
-
[16]
Tengfei Wang and Xiufen Zou. Dynamic analysis of a drug resistance evolution model with nonlinear immune response.Math Biosci, 374:109239, August 2024
work page 2024
-
[17]
Jeffrey West, Li You, Jingsong Zhang, Robert A. Gatenby, Joel S. Brown, Paul K. Newton, and Alexander R.A. Anderson. Towards multidrug adaptive therapy.Cancer Res, 80(7):1578–1589, April 2020. 31
work page 2020
-
[18]
F. J. Burns and I. F. Tannock. On the existence of a G0 -phase in the cell cycle.Cell Tissue Kinet, 3(4):321–334, 1970
work page 1970
-
[19]
Jinzhi Lei. A general mathematical framework for understanding the behavior of hetero- geneous stem cell regeneration.J Theor Biol, 492:110196, May 2020
work page 2020
-
[20]
Samuel Bernard, Jacques B´ elair, and Micahel C. Mackey. Oscillations in cyclical neu- tropenia: new evidence based on mathematical modeling.J Theor Biol, 223(3):283–298, May 2003
work page 2003
-
[21]
Michael C Mackey. Unified hypthesis for the origin of aplasgtic anemia and periodic hematopoiesis.Blood, 51(5):941–956, 1978
work page 1978
-
[22]
Jinzhi Lei. Evolutionary dynamics of cancer: from epigenetic regulation to cell population dynamics – mathematical model framework, applications, and open problems.Sci China Math, 63(3):411–424, 2020
work page 2020
-
[23]
Can Zhang, Changrong Shao, Xiaopei Jiao, Yue Bai, Miao Li, Hanping Shi, Jinzhi Lei, and Xiaosong Zhong. Individual cell-based modeling of tumor cell plasticity-induced immune escape after CAR-T therapy.Comput Syst Oncol, 1:e21029, 2021
work page 2021
-
[24]
Rongsheng Huang, Qiaojun Situ, and Jinzhi Lei. Dynamics of cell-type transition medi- ated by epigenetic modifications.J Theor Biol, 577:111664, Jan 2024
work page 2024
-
[25]
Yakun Li, Xiyin Liang, and Jinzhi Lei. Integrating gene regulatory network dynamics with heterogeneous stem cell regeneration.CSIAM Trans Life Sci, 1(2):320–353, 2025
work page 2025
-
[26]
Jeffrey J Quinn, Matthew G Jones, Ross A Okimoto, Shigeki Nanjo, Michelle M Chan, Nir Yosef, Trever G Bivona, and Jonathan S Weissman. Single-cell lineages reveal the rates, routes, and drivers of metastasis in cancer xenografts.Science, 371(6532), Feb 2021
work page 2021
-
[27]
Annina Denoth-Lippuner, Baptiste N Jaeger, Tong Liang, Lars N Royall, Stefanie E Chie, Kilian Buthey, Diana Machado, Vladislav I Korobeynyk, Merit Kruse, Clara M Munz, Alexander Gerbaulet, Benjamin D Simons, and Sebastian Jessberger. Visualiza- tion of individual cell division history in complex tissues using iCOUNT.Cell Stem Cell, 28(11):2020–2034, Nov 2021
work page 2020
-
[28]
Oscillatory dynamics of heterogeneous stem cell regeneration
Xiyin Liang and Jinzhi Lei. Oscillatory dynamics of heterogeneous stem cell regeneration. Com Appl Math Comput, 6:431–453, 2024
work page 2024
-
[29]
Shun Wang, Jinzhi Lei, Xiufen Zou, and Suoqin Jin. Integrating multiscale mathemat- ical modeling and multidimensional data reveals the effects of epigenetic instabiility on acquired resistance in cancer.PLoS Comp Biol, 21(2):e1012815, 2025
work page 2025
-
[30]
Population ecology issues in tumor growth.Cancer Res, 51:2542– 2547, 1991
Robert A Gatenby. Population ecology issues in tumor growth.Cancer Res, 51:2542– 2547, 1991
work page 1991
-
[31]
D Ravindran Menon, S Das, C Krepler, A Vultur, B Rinner, S Schauer, K Kashofer, K Wagner, G Zhang, E Bonyadi Rad, N K Haass, H P Soyer, B Gabrielli, R Somasun- daram, G Hoefler, M Herlyn, and H Schaider. A stress-induced early innate response causes multidrug tolerance in melanoma.Oncogene, 34(34):4448–4459, 2015
work page 2015
-
[32]
Brian B Liau, Cem Sievers, Laura K Donohue, Shawn M Gillespie, William A Flavahan, Tyler E Miller, Andrew S Venteicher, Christine H Hebert, Christopher D Carey, Scott J Rodig, Sarah J Shareef, Fadi J Najm, Peter van Galen, Hiroaki Wakimoto, Daniel P Cahill, Jeremy N Rich, Jon C Aster, Mario L Suv` a, Anoop P Patel, and Bradley E Bernstein. Adaptive chroma...
work page 2017
-
[33]
Sreenath V Sharma, Diana Y Lee, Bihua Li, Margaret P Quinlan, Fumiyuki Takahashi, Shyamala Maheswaran, Ultan McDermott, Nancy Azizian, Lee Zou, Michael A Fis- chbach, Kwok-Kin Wong, Kathleyn Brandstetter, Ben Wittner, Sridhar Ramaswamy, Marie Classon, and Jeff Settleman. A chromatin-mediated reversible drug-tolerant state in cancer cell subpopulations.Cel...
work page 2009
-
[34]
Mustafa Asowed, Nils O Elander, Linn Pettersson, Maria Ekholm, and Dimitrios Pa- pantoniou. Activity and safety of kees - an oral multi-drug chemo-hormonal metronomic combination regimen in metastatic castration-resistant prostate cancer.BMC Cancer, 23(1), April 2023. 33
work page 2023
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.